Zebrafish: A window into myeloid cell development, disease, and defense
Gaofei Li

Jiawen Lao
Jialong Deng
Shengbo Liu
Rui Liang
Yiyue Zhang
*
*Correspondence to:
Yiyue Zhang, The Innovation Centre of Ministry of Education for Development and Diseases, Division of Cell, Developmental and Integrative Biology, School of Medicine, South China University of Technology, Guangzhou 510006, Guangdong, China.
E-mail: mczhangyy@scut.edu.cn.
Myeloid Cells. 2026;1:202505. 10.70401/mc.2026.0002
Received: March 30, 2026Accepted: May 18, 2026Published: May 18, 2026
Abstract
Myeloid cells play key roles in innate and adaptive immunity. Studying their development and function helps reveal new roles and pathogenic mechanisms of related diseases. Zebrafish, a classical model organism, offers various advantages for studying myeloid cell biology. In this review, we focus on recent advances in myeloid cell development and function using the zebrafish model. The pattern and regulation of myelopoiesis and related disorders, including congenital myeloid cell insufficiency and myeloid malignancies, have been revealed, along with related drugs. Regarding function, we highlight myeloid cells, especially macrophages, in homeostasis and regeneration. The bidirectional interactions between myeloid cells and pathogens in zebrafish also provide valuable insights into the mechanisms of infectious diseases. In summary, we highlight the unique contributions of the zebrafish model, demonstrate its complementary roles in basic research and clinical applications relative to mammalian systems, and look forward to more exciting discoveries using this model in the future.
Keywords
Myeloid cell, zebrafish, myelopoiesis, leukemia, immune function, regeneration
1. Introduction
Myeloid cells are important components of the immune system, including granulocytes (neutrophils, eosinophils, basophils, and mast cells) and other myeloid cells (monocytes/macrophages and dendritic cells (DCs))[1]. They are all derived from hematopoietic stem cells (HSCs) and multipotent progenitors, then bifurcated into different lineages during cell fate determination and differentiation[2]. After myelopoiesis, mature myeloid cells are found in various tissues and organs, where they play complex roles in both homeostasis and diseases[1,3].
Although most studies on myeloid cells were conducted in mammals, zebrafish, with its unique advantages, has also contributed important findings to the field. In this review, we focus on the progress in understanding myeloid cell development and function using the zebrafish model. These studies reveal myeloid cell biology from a unique perspective and provide a strong complement to mammalian research.
2. Why Zebrafish?
As a typical teleost model organism, zebrafish possesses several advantages for studying myeloid cells. First, zebrafish share high genetic conservation with humans. Over 80% of genes associated with human diseases have definite zebrafish orthologs[4], making zebrafish suitable for modeling many human diseases[5]. Furthermore, zebrafish are characterized by easy accessibility for genetic manipulation. Gene knockout is widely used in zebrafish[6], and advances in base editing technologies[7-10] have enabled more precise modeling of human inherited diseases. Although the low efficiency of homologous recombination in the fast-developing zebrafish embryo has hindered the generation of conditional knockout (CKO) models[11], recent approaches have largely overcome this limit[11-13], and neutrophil-specific CKO has been achieved[14]. In addition, rapid gene knockdown through morpholino[15] or clustered regularly interspaced short palindromic repeats (CRISPR)/Cas9-based disruption[16-18] also makes zebrafish suitable for rapid target gene evaluation. Apart from these reverse genetic approaches, large-scale forward genetic screens have also been applied in zebrafish to identify new genes associated with specific phenotypes[19,20]. Taken together, based on genome conservation and genetic accessibility, zebrafish are an ideal model for investigating gene function and modeling human diseases.
Second, zebrafish embryos are transparent and develop externally from the 1-cell stage[19], making them an outstanding model for developmental biology. Based on the Tol2 transposon system, short promoter-gene elements[21] or large bacterial artificial chromosomes (BAC)[22] can be randomly integrated into the zebrafish genome to generate transgenic lines. This methodology enables target gene overexpression or cell ablation based on metronidazole/nitroreductase[23] or diphtheria toxin A[24] systems, facilitating studies of gene and cell function. More importantly, using myeloid-specific promoters to drive fluorescence expression allows specific labeling of myeloid cells. To date, a series of reporter lines have been generated (Table 1), enabling dynamic studying of myeloid cell development, behavior, and function in the transparent larvae. For instance, lattice lightsheet microscopy technology has revealed detailed neutrophil and macrophage behaviors during homeostasis on the subcellular level[25].
Table 1. Reported transgenic zebrafish lines with myeloid cell labeling.
| Cell type | Promoter | Transgenic line | Brief description | Reference |
| Myeloid cells | coro1a | Tg(coro1a:eGFP) | All myeloid cells are labeled with eGFP. | [26] |
| Myeloid cells | coro1a | Tg(coro1a:loxP-DsRedx-loxP-GFP) | All myeloid cells are labeled with DsRedx but can be switched to GFP+ upon cre/loxP recombination. | [27] |
| Macrophages and DCs | mpeg1 | Tg(mpeg1:eGFP) | Macrophages and DCs are labeled with eGFP. | [28,29] |
| Macrophages and DCs | mpeg1 | Tg(mpeg1:mCherry) | Macrophages and DCs are labeled with mCherry. | [29] |
| Macrophages and DCs | mpeg1 | Tg(mpeg1:loxP-DsRedx-loxP-GFP) | Macrophages and DCs are labeled with DsRedx but can be switched to GFP+ upon cre/loxP recombination. | [30] |
| Macrophages and DCs | mpeg1 | Tg(mpeg1:Dendra2) | Macrophages and DCs undergo photoconversion from green to red fluorescent states upon exposure to a 405 nm laser. | [31] |
| Macrophages and DCs | mpeg1 | Tg(mpeg1:Kaede) | Macrophages and DCs undergo photoconversion from green to red fluorescent states upon exposure to a 405 nm laser. | [32] |
| Macrophages and DCs | mhc2dab | Tg(mhc2dab:GFP) | The GFP+ cells are predominantly macrophages and DCs. | [33] |
| Macrophages | mfap4 | Tg(mfap4:mCherry) | Macrophages are labeled with mCherry. | [34] |
| Macrophages | c1qb | Tg(c1qb:EGFP) | Macrophages are labeled with eGFP. | [35] |
| Conventional DCs | ccl35.1 | TgBAC(ccl35.1:EGFP) | Conventional DCs are labeled with eGFP. | [35] |
| Pro-inflammatory macrophages | lgals3bpb | Tg(lgals3bpb:EGFP) | Pro-inflammatory macrophages are labeled with eGFP. | [35] |
| Pro-remodeling macrophages | ccl34a.4 | TgBAC(ccl34a.4:EGFP) | Pro-remodeling macrophages are labeled with eGFP. | [35] |
| Microglia | ccl34b.1 | TgBAC(ccl34b.1:EGFP) | Microglia are labeled with eGFP. | [36] |
| Neutrophils | mpx | Tg(mpx:GFP) | Neutrophils are labeled with GFP. | [37] |
| Neutrophils | mpx | Tg(mpx:Dendra2) | Neutrophils undergo photoconversion from green to red fluorescence upon exposure to a 405 nm laser. | [38] |
| Neutrophils | lyz | Tg(lyz:GFP) | Neutrophils are labeled with GFP. | [39,40] |
| Neutrophils | lyz | Tg(lyz:CFP) | Neutrophils are labeled with CFP. | [41] |
| Neutrophils | lyz | Tg(lyz:DsRed) | Neutrophils are labeled with DsRed. | [40] |
| Mature neutrophils | mmp9 | TgBAC(mmp9:Citrine) | Mature neutrophils are labeled with Citrine. | [41] |
| Neutrophils and mast cells | cpa5 | Tg(cpa5:GFP) | Around 80% of GFP+ cells are neutrophils, while 20% are mast cell-like cells. | [42] |
| Eosinophils | gata2a | Tg(gata2a:eGFP) | The eGFP+ cells in adult hematopoietic tissues are eosinophils. | [43] |
| Eosinophils | eslec | Tg(eslec:eGFP) | Eosinophils are labeled with eGFP. | [44] |
| Eosinophils | eslec | Tg(eslec:DsRed) | Eosinophils are labeled with DsRed. | [44] |
| Eosinophils | eslec | TgKI(eslec-tdTomato) | Eosinophils are labeled with tdTomato, which is fused with Eslec to show Eslec localization. | [45] |
eGFP/EGFP: enhanced green fluorescent protein; GFP: green fluorescent protein; CFP: cyan fluorescent protein; DCs: dendritic cells.
Finally, each pair of adult zebrafish can produce ~200 embryos per week. This high fecundity makes zebrafish a high-throughput in vivo model for drug screening[5,46]. Additionally, because zebrafish are continuously exposed to the aquatic environment, they are particularly valuable for toxicology studies using environmental pollutants or toxins[46]. Collectively, these advantages make zebrafish a powerful model for studying myeloid cells across multiple fields, including developmental biology, immunology, drug discovery, and toxicology.
3. Zebrafish Myeloid Cells
Most myeloid cell types are present in zebrafish, with the exception of basophils[47]. The morphology and distribution of myeloid cells during zebrafish early development have been characterized for decades[47,48]. In recent years, single-cell RNA sequencing (scRNA-Seq) atlases of larval and adult zebrafish have further demonstrated the distribution and gene expression profiles of myeloid cells on the whole-organism level[49-52].
For macrophages and DCs, recent scRNA-Seq studies revealed their presence in various organs including the kidney, thymus, epidermis, gill, intestine, liver, heart, and brain[35,53,54]. Macrophages can be further distinguished into pro-inflammatory or pro-remodeling subsets[35], while conventional and plasmacytoid DC subpopulations have also been found in zebrafish DCs[35,54]. These findings highlight the overall conservation of macrophages and DCs between zebrafish and mammals.
Among granulocytes, neutrophils are the most abundant and well-characterized type. Comparative transcriptomics has confirmed high molecular similarities among zebrafish, mouse, and human neutrophils[41]. Eosinophils are found in various adult tissues[43,44], and their lineage features from progenitors to mature cells have been deciphered by scRNA-Seq[44]. Mast cells remain the least investigated myeloid cell type in zebrafish. Their presence has been confirmed by Toluidine blue staining, but their specific markers and lineage features remain unclear[42,55], warranting further investigation.
Another zebrafish myeloid-like cell type, metaphocytes, transfer soluble antigens to conventional Langerhans cells and are essential for barrier immunity[56,57]. Although the metaphocytes are mpeg1-positive, they are derived from ectoderm and thus are not considered typical myeloid cells[56]. Accordingly, this review will not focus on this cell type. Taken together, most myeloid cell types have been characterized in zebrafish and show conservation with their mammalian counterparts.
4. Zebrafish Myelopoiesis
4.1 Myeloid progenitor development and regulation
Similar to mammals, zebrafish hematopoiesis occurs in three successive waves (Figure 1). Primitive myelopoiesis begins around 11 hours post fertilization in the rostral blood island (RBI)[58]. The intermediate wave originates from endothelium-derived hematopoietic progenitors (EHPs) in the posterior blood island (PBI) and aorta-gonad-mesonephros (AGM) regions[59]. The definitive wave originates from HSCs, which are derived from hemogenic endothelium through endothelial-to-hematopoietic transition[60,61]. HSCs then migrate to the caudal hematopoietic tissue (CHT) for expansion and subsequently colonize the kidney marrow (KM) (the functional equivalent of mammalian bone marrow), giving rise to all adult myeloid cells. Based on the conservation of the three waves of hematopoiesis, the zebrafish model, due to its unique advantages in developmental biology, is particularly suitable for studying primitive and intermediate myelopoiesis.
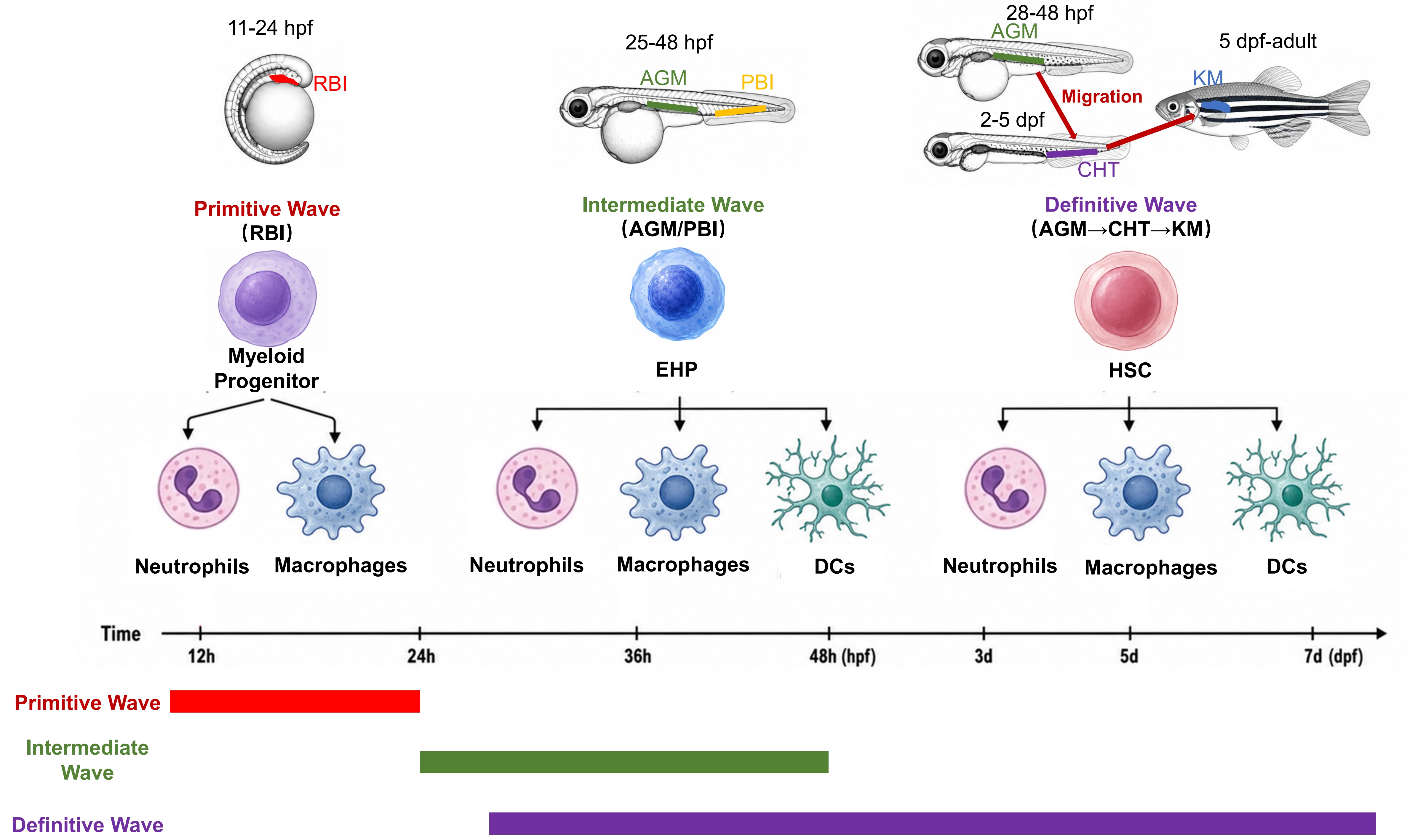
Figure 1. Schematic of zebrafish myelopoiesis. The primitive wave originates from RBI and generates primitive neutrophils and macrophages. The intermediate wave originates from both AGM and PBI and generates neutrophils, macrophages and DCs. The definitive wave originates from AGM, and the HSCs migrate to CHT and finally KM, generating neutrophils, macrophages and DCs. The origin of eosinophils and mast cells remain unknown and thus are not included in this schematic. RBI: rostral blood island; AGM: aorta-gonad-mesonephros; PBI: posterior blood island; DCs: dendritic cells; HSCs: hematopoietic stem cells; CHT: caudal hematopoietic tissue; KM: kidney marrow.
Transcription factors (TFs) play key roles in the regulation of myelopoiesis (Table 2). Cebpα acts cooperatively with Runx1 and is essential for myeloid progenitors in producing mature myeloid cells[62]. Suppression of Pu.1 also results in severe defects of all myeloid cells[63]. For other regulators, Alk8 is required for pu.1 expression and primitive myelopoiesis[77]. In addition, the orphan enzyme Peds1 inhibits myeloid cell apoptosis to promote myelopoiesis[78].
Table 2. Reported TFs in the development of myeloid cell lineages.
| TF | Functions and related references |
| Cebpα | Essential for myelopoiesis[62] |
| Pu.1 | Essential for myelopoiesis[63] |
| Gata2a | Inducing primitive myelopoiesis[64] |
| Runx1 | Promoting neutrophil fate and inhibiting macrophage fate[65]; facilitating neutrophil maturation[66] |
| Irf8 | Promoting macrophage fate over neutrophil fate[67,68] |
| Irf2bp2b | Promoting neutrophil fate over macrophage fate[69] |
| Ikzf1 | Promoting neutrophil fate over macrophage fate[68] |
| cMyb | Promoting neutrophil fate over macrophage fate[68]; facilitating neutrophil maturation[39,66] |
| Spi-b | Inducing macrophage development[28] |
| Elf1 | Inducing macrophage development[70] |
| Mafbb | Inhibiting osteoclast differentiation[71] |
| Zeb2a | Essential for microglia development and suppressing brain DC development[72] |
| Batf3 | Required for brain conventional DC development[72] |
| Gfi1aa | Inhibiting neutrophil differentiation[73] |
| Tead1a | Promoting neutrophilopoiesis[74] |
| Znf687 | Suppressing neutrophilopoiesis[75] |
| Cebp1 | Facilitating neutrophil maturation[39]; inhibiting eosinophil lineage fate[44] |
| Gata2b | Inducing neutrophilopoiesis[64] |
| Cebpβ | Essential for emergency granulopoiesis[76]; promoting eosinophil lineage fate[44] |
TF: transcription factor; DC: dendritic cell.
The mechanisms governing fate choice among myeloid lineages have also been investigated in zebrafish. Primitive myelopoiesis cell fate is regulated by blf and the drl cluster, whose depletion leads to excessive macrophage development at the expense of neutrophils[79]. Interferon regulatory factor-8 (Irf8) is another key factor that promotes macrophage fate over neutrophil fate during primitive myelopoiesis[69]. The Ikzf1-cMyb complex targets the lncRNA irf8-2, which further modulates Irf8 levels via Zfp36l1, thereby affecting the neutrophil versus macrophage commitment[70]. In addition, Irf2bp2b, acting downstream of Cebpα, represses pu.1 to induce neutrophil lineage bias at the expense of macrophages[71]. Collectively, these studies in zebrafish have identified multiple factors that shape myeloid lineages by regulating progenitor differentiation and fate determination.
4.2 Macrophage and DC development and regulation
For the regulation of macrophage development, primitive macrophages derived from the RBI are regulated through a hierarchical cascade in which Pu.1 acts upstream of Spi-b, and together they control irf8 expression to drive macrophage differentiation[28]. In contrast, macrophages originating from the AGM are regulated by Pu.1 and Spi-b in a parallel manner, which converges on Irf8 to induce macrophage development[28]. In addition, Pu.1 and Irf8 also directly induce granulin (grna) expression to promote macrophage development[80]. Other factors such as tripartite motif-containing protein 33 (Trim33)[81], E74-like factor 1 (Elf1)[72], colony-stimulating factor 1 receptor (Csf1r)[82], and the mineralocorticoid receptor[83] have also been shown to affect macrophage development.
Tissue-resident macrophages are present in most tissues, such as Kupffer cells in the liver, osteoclasts in the bone, Langerhans cells in the skin, and microglia in the central nervous system (CNS)[84]. For their development, Xu et al. reported that microglial precursors colonize the larval zebrafish midbrain in a circulation-independent manner, and this process is promoted by apoptotic neuronal cells[85]. Lineage tracing has exhibited that embryonic microglia arise from RBI and depend on Pu.1 and IL34[27,86], whereas adult microglia arise from AGM and depend on Runx1 and cMyb[27,32]. However, adult mouse microglia mainly originate from yolk sac-derived erythron-myeloid precursors[87], and the difference may be due to species variation or distinct fate-mapping approaches[30]. Pu.1 and Spi-b regulate microglia maintenance in a dosage-dependent manner, which effect is conserved in mice[88]. In addition, Zeb2a is also found essential for microglia development[74]. For osteoclasts, Pu.1[89], Fms[89], and Mafbb[73] play key roles in their differentiation. The colonization and differentiation of macrophages in other tissues and the regulation mechanisms await further investigation in zebrafish, which may reveal how macrophages adapt to the tissue niches from a dynamic perspective.
In larval zebrafish, the initial transient wave of DCs, derived from EHPs of the PBI region, emerges since 3 days post fertilization (dpf) only in the thymus, whereas the subsequent definitive wave of DCs originates from HSCs and persist into adulthood[90]. Regarding developmental regulation, Lin et al. demonstrated that Flt3 is dispensable for larval DCs but required for the survival of adult DCs, especially those residing in the brain[90]. The Id2a-Batf3 axis is also required for brain DC development, while Zeb2a inhibits Id2a to suppress this process[74]. Research on DCs in zebrafish is still emerging and holds promise for providing further insights into DC biology.
4.3 Granulocyte development and regulation
In zebrafish, neutrophilopoiesis and its underlying mechanisms have been studied for years[91], and here we will focus on recent advances. Neutrophils were found to be derived from the RBI or CHT region, with their features characterized by scRNA-Seq[92]. TFs play indispensable roles in neutrophil regulation. Tead1a activates the Tead1a/Yes-associated protein (YAP)-Notch1-Spi1/Cebpα axis to promote neutrophilopoiesis[76]. cMyb cooperates with Runx1[68] and Cebp1[39] to facilitate neutrophil maturation. Gfi1aa-deficient zebrafish exhibits accumulation of immature neutrophils, which is due to abolished Gfi1aa-Lsd1 inhibition on cebpa[75]. Irf2bp2a and Gfi1aa suppress each other to form a negative feedback loop in regulating neutrophilopoiesis[93]. Znf687 negatively regulates neutrophil development by recruiting the Brd4-Smrt complex to repress gfi1aa[77]. Beyond TFs, vesicle dynamic defects (vps18 mutation)[94], carboxypeptidase defects (cpa1 and cpa5 mutation)[42], and cytokine signaling dysregulation[95] also affect neutrophilopoiesis.
Eosinophils emerge since 5 dpf and become relatively abundant by 7 dpf[44]. In both larval and adult zebrafish, eosinophils are mainly found in the kidney, intestine, and peritoneal fluid[43,44]. A transcriptional axis formed by Cebp1 and Cebpβ regulates eosinophilopoiesis by affecting lineage fate determination and differentiation[44]. The recently established eosinophil reporter lines in zebrafish may help reveal more regulatory mechanisms, and together with findings in the mammalian system, will enhance our understanding of eosinophilopoiesis.
4.4 Myelopoiesis disorder models and drug development
Congenital myeloid cell insufficiency results in severe immune deficiencies. Mutation of dnajc21 in zebrafish models Shwachman-Diamond syndrome with a neutropenia phenotype, which can be rescued by TP53[96]. Deficiency in zebrafish srp54, srp19, or srpra induces congenital neutropenia (CN), highlighting the indispensable roles of the signal recognition particle receptor (SRP) ribonucleoprotein complex in neutrophils[97,98]. Knockdown of zebrafish hax1 also induces CN, which can be reversed by granulocyte-colony stimulating factor (G-CSF) or flavopiridol[99,100]. Flavopiridol additionally restores CN phenotypes caused by jagn1b knockout or cebpa knockdown[100]. Mutation of copz1 also results in CN, which can be restored by the small molecule HIF1α stabilizer IOX2[101]. In summary, congenital myeloid disorders have been modeled in zebrafish, and relevant therapeutic agents have been identified.
Myeloid malignancies, including myelodysplastic syndrome (MDS), myeloproliferative neoplasms (MPN), acute myeloid leukemia (AML), and chronic myelomonocytic leukemia (CMML), are characterized by abnormal proliferation and differentiation of myeloid progenitors. Numerous zebrafish models of these malignancies have been established, and effective drugs for disease relief have been identified[102]. Here, we focus on recent advances. Loss-of-function of tet2 with further induction of stress hematopoiesis can lead to a proliferative pre-leukemic phenotype[103]. C-terminal truncated mutation of asxl1 induces neutrophilic dysplasia, which is a foundation for myeloid malignancies[104], while further transgenic overexpression of SRSF2P95H in asxl1 mutants recapitulates CMML phenotypes[105]. Transgenic IDH2R172K and IDH2R140Q zebrafish models exhibit AML features, which can be ameliorated by quizartinib and enasidenib[106]. A knockin mutation of Shp2D61G results in MPN, which can be rescued by dexamethasone[107]. Xu et al. reported that cMyb hyperactivation and Pu.1 deficiency synergistically drive the progression of MDS and secondary AML, a phenotype effectively relieved by cytarabine and all-trans retinoic acid[108]. In summary, these studies not only model human myeloid malignancies, but also suggest new therapeutic strategies for potential clinical translation.
5. Zebrafish Myeloid Cell Function
5.1 Myeloid cells in homeostasis
First, we discuss myeloid functions in the absence of wounds or infection. Several studies have suggested the roles of primitive macrophages in establishing definitive hematopoiesis. Depletion of primitive macrophages using liposome encapsulated clodronate or nitroreductase reduces hematopoietic stem/progenitor cell (HSPC) colonization in the CHT[109]. However, cebpa mutants, which lack all primitive macrophages, exhibit normal HSPC numbers and CHT colonization[110]. This discrepancy may arise because cell death-dependent depletion of macrophages leads to stimuli that affect hematopoiesis. Vcam-1-positive macrophages interact with HSPCs to promote their homing and retention to a vascular niche in the CHT[111]. A more recent study reported that CHT macrophages interact with nascent HSPCs and monitor their quality, resulting in either engulfment and elimination of HSPCs or their expansion[112]. Therefore, primitive macrophages may not affect total HSPC numbers but control HSPC clonality instead.
For other myeloid cell functions, microglia perform phagocytosis to facilitate tissue clearance, whereas brain DCs exert regulatory functions by releasing immune regulators[36]. A type I interferon-responsive microglial subset that engulfs neurons is required for normal cortical development and sensorimotor function[113]. In addition, both neutrophils and macrophages contact thyrocytes and promote their migration during thyroid follicle formation[114]. Hu et al. recently revealed a role of macrophages in regulating systemic iron homeostasis and atherosclerosis[115]. Osteoclasts sculpt craniofacial form to permit sensorineural patterning[116]. On the other hand, macrophages promote ovarian failure and masculinization in zebrafish[117]. These studies underscore the diverse functions of zebrafish myeloid cells, particularly macrophages, in homeostasis.
5.2 Myeloid cells in pathogen clearance
Leveraging transparency and myeloid cell labeling, myeloid cell behaviors during infection can be dynamically monitored in zebrafish. Upon wounds or Escherichia coli infection, unpolarized macrophages migrate to the inflammation sites and polarize into M1 macrophages[118]. Long-term tracing has exhibited that the same macrophage population eventually converts into M2 macrophages to resolve inflammation[118]. Upon wounds or Staphylococcus aureus infection, neutrophils swarm toward early recruited pioneer neutrophils, which violently release neutrophil extracellular traps[119].
During bacterial infection, both neutrophils and macrophages can phagocytose bacteria such as Streptococcus[31,120] and Salmonella[40,121] to protect zebrafish survival. Shigella induces reprogramming of zebrafish neutrophils and pro-inflammatory macrophages, enhancing the antimicrobial activity of trained neutrophils and macrophages[122,123]. However, in some cases, bacteria take advantage of macrophages to evade clearance. For instance, the switch from pro-inflammatory macrophages into anti-inflammatory macrophages provides a permissive niche for persistent Salmonella typhimurium[124]. During Mycobacterium leprae infection, the infected macrophages even produce reactive oxygen species (ROS) to damage axons, initiating nerve damage in leprosy[125].
Among the bacteria, Mycobacterium tuberculosis is one of the most threatening pathogens to humans, and the zebrafish Mycobacterium marinum infection model, which recapitulates granuloma formation, provides an important tool for studying its pathogenesis[126]. Luo et al. reported that a specific subset of macrophages within a developing granuloma helps control M. marinum growth[127]. Neutrophils are subsequently recruited to granulomas by dying infected macrophages and phagocytose and kill the internalized mycobacteria[128]. Eosinophils are also present in the necrotic granulomas of zebrafish, potentially exerting protective roles in host survival[129]. However, M. marinum can reprogram macrophages to induce granuloma formation, thereby limiting immune cell access[130], and can also hijack macrophages to achieve dissemination[131]. In a recent study, Viswanathan et al. reported that neutrophils can also facilitate mycobacterial survival within granulomas[132]. Together, these findings highlight the complex roles of myeloid cells in mycobacterial infection, which may vary by infection stage and myeloid cell heterogeneity (Figure 2).
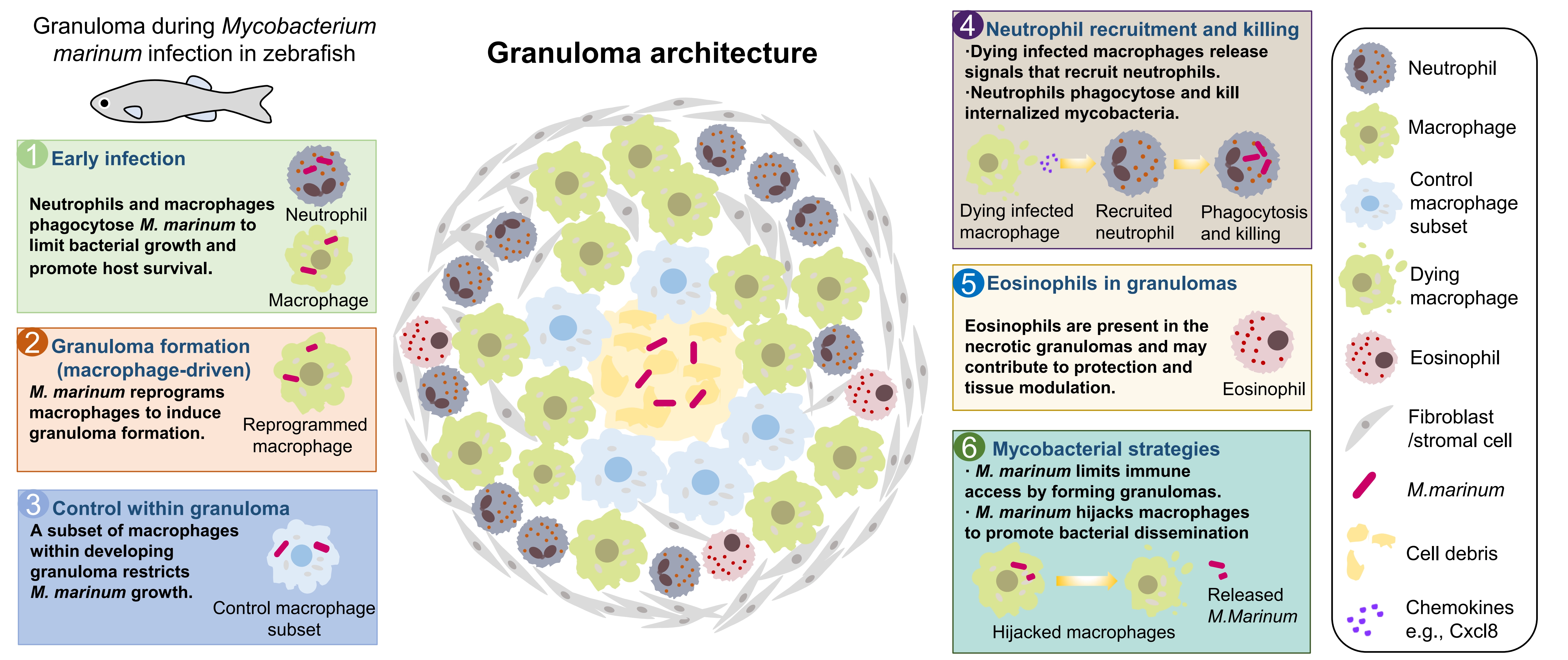
During viral infection, a scRNA-Seq analysis revealed the myeloid cell landscapes in the spleen following spring viremia of carp virus (SVCV) infection, and showed that vaccination with inactivated SVCV induces trained immunity responses of neutrophils and M1 macrophages[133]. During the infection of human norovirus (HuNoV), macrophages, but not neutrophils, are infected and identified as permissive cells for HuNoV replication[134].
During fungal infection, macrophages can phagocytose Cryptococcus neoformans to protect zebrafish larvae[135]. However, phagocytosis of Talaromyces marneffei by macrophages protects the fungi from neutrophil fungicidal activity[136]. Neutrophil deficiency renders zebrafish embryos more susceptible to Aspergillus fumigatus, whereas stimulating autophagy responses improves host survival[137]. During Aspergillus fumigatus infection, macrophages inhibit spore germination to promote fungal persistence, while neutrophils kill fast-germinating fungi[138], illustrating the complex roles of neutrophils and macrophages in infections with rapid growing or dormitive fungi.
During parasitic infection, Schistosoma mansoni egg can induce macrophage recruitment, contributing to granuloma formation[139]. In Trypanosoma carassii infection, macrophages exhibit stronger responses than neutrophils, and a unique foamy macrophage subset with strong pro-inflammatory features emerges[140]. Although both neutrophils and macrophages are recruited to the infected Ichthyophthirius multifiliis, the vigorous rotation of parasites largely prevents clearance by these myeloid cells[141], and this phenomenon is also found in T. carassii infection[140]. Eosinophils degranulate in response to Heligmosomoides polygyrus extract and are recruited to the intestine upon Pseudocapillaria tomentosa infection[43]. Therefore, current studies mainly focus on myeloid cell responses during parasite infection, but their precise functions remain elusive, especially for eosinophils, the professional anti-parasitic cells in mammals.
In some cases, pathogens induce myeloid cell death to escape from innate immunity. For instance, Salmonella typhimurium utilizes programmed cell death pathways to kill neutrophils and macrophages[142]. In tuberculous granuloma, mammalian target of rapamycin (mTOR) protects macrophages from mycobacterium-induced cell death[143], whereas tumor necrosis factor (TNF) facilitates macrophage cell death, an effect that can be prevented by metformin[144]. Treponema pallidum promotes microglia apoptosis to evade clearance in the CNS, contributing to neurosyphilis[145]. In summary, studies using the zebrafish model have reported bidirectional interactions between myeloid cells and pathogens, providing valuable insights into the mechanisms of infectious diseases.
5.3 Myeloid cells in inflammatory disease models
Many immune-mediated inflammatory diseases have been modeled in zebrafish[146,147]. Most studies focused on inflammatory bowel disease (IBD), which is a chronic inflammatory disorder of the gastrointestinal tract[148,149]. During colitis, neutrophils can disrupt the intestinal barrier via the IL-22/transforming growth factor β (TGF-β)/Mmp9 axis[150]. Lemairamin inhibits the recruitment of zebrafish neutrophils to the intestinal injury, thus exerting its anti-inflammatory effect in IBD[151]. On the other hand, the recruitment of macrophages to the intestine is required for intestinal inflammatory lymphangiogenesis[152]. In addition to IBD models, evidence from a systemic inflammation zebrafish model demonstrates that chronic macrophage activation impairs muscle regeneration by suppressing repair programs[153]. In summary, the roles of myeloid cells in other inflammatory disease models warrant further exploration and may provide new insights into pathogenic mechanisms.
5.4 Myeloid cells in wound healing and regeneration
Zebrafish have a strong regeneration capability that can fully regenerate various organs, making it an outstanding model for studying regeneration mechanisms[154]. A classical regeneration model is tail fin amputation in larval zebrafish, with wound recovery occurring within several days[155]. After tail fin transection, both neutrophils and macrophages migrate toward the injury site[29,37], with neutrophils responding much faster than macrophages[29]. Long-term tracking has revealed bidirectional traffic of neutrophils between the wound and vasculature, and the neutrophils rarely undergo apoptosis[38]. Neutrophils serve as primary scavengers during the inflammation stage, but are dispensable for fin regeneration[26]. Macrophages, recruited by calcium and ROS[156], participate in neutrophil phagocytosis and are essential for inflammation resolution and fin regeneration[26,29].
Unlike mammals, zebrafish undergo neurogenesis after CNS injury to support regeneration. Zebrafish microglia are responsible for phagocytosis of dead neurons[157,158]. Real-time imaging revealed that dead neuron signals spread in the form of calcium waves, which recruit microglia to injury sites[159]. In an oligodendrocyte depletion model, DCs undergo stage-specific morphological remodeling, suggesting active involvement in myelin repair[160]. Following spinal cord injury, neutrophils are rapidly recruited but not essential for axonal regrowth[161]. Conversely, a pro-regenerative macrophage subset produces Tnf to influence ependymo-radial glia progenitor cells, thereby playing vital roles in spinal cord neurogenesis[161,162]. By means of live imaging, Denans et al. demonstrated an M1-to-M2 switch and anti-inflammatory pathways in macrophages during hair cell regeneration in the zebrafish lateral line[163]. Additional studies have highlighted key roles of retinal microglia in retinal regeneration[164,165]. Collectively, these studies underscore the importance of macrophages in zebrafish neuronal regeneration, offering insights with potential application for mammalian systems.
Another remarkable capability of zebrafish is the ability to fully regenerate the heart, whereas cardiac injury in mammals leads to scarring[166]. After cardiac injury, neutrophils, macrophages, and eosinophils are recruited to the heart[167]. ScRNA-Seq analysis has revealed the heterogeneity and features of macrophages and neutrophils during cardiac regeneration[168]. Rapid accumulation of neutrophils is important for clearing dead cells and debris[169]. Among macrophages, tnfa-positive pro-inflammatory macrophages promote scar deposition[167,170], whereas tnfa-negative anti-inflammatory macrophages facilitate scar resolution[167]. In addition, neutrophils are essential for the regeneration of the epicardium[171], and epicardial Vegfaa signaling recruits macrophages to the epicardial-myocardial niche to induce cardiomyocyte proliferation[172]. The above studies demonstrate the key roles of neutrophils and macrophages during zebrafish cardiac regeneration.
6. Conclusion
In this review, we highlight recent advances in the study of myeloid cells using the zebrafish model (Figure 3). Key insights have emerged regarding spatiotemporal developmental patterns, origins, and associated disorders of myelopoiesis in zebrafish. Furthermore, the functional roles of myeloid cells in homeostasis, pathogen clearance, and tissue regeneration are discussed. Collectively, these studies underscore the unique advantages of the zebrafish model for dynamically investigating myeloid cell biology.
Figure 3. Schematic of zebrafish model in studying myeloid cell biology. DC: dendritic cell.
The zebrafish model holds promise for uncovering more myeloid cell functions. Leveraging the regenerative functions of zebrafish myeloid cells may inform strategies to promote mammalian organ regeneration. In the era of artificial intelligence, machine learning could enable deeper and more automated analyses of myeloid cell behaviors. Such approaches would capitalize on the imaging advantages of zebrafish to explore myeloid cell dynamics under various genetic perturbations, disease models, or drug treatments, offering new perspectives on myeloid cell functions.
As the functions of zebrafish myeloid cells become increasingly defined, a central question is how to leverage these functions and their underlying mechanisms. Drug development may be a key avenue. Owing to its high fecundity, zebrafish is the only vertebrate model organism suitable for large-scale drug screening. In this context, model accuracy is critical, and zebrafish offer several advantages over mice. For example, mice are nocturnal with rod-dominant retinas, whereas zebrafish have cone-dominant retinas similar to humans[173]. In addition, the zebrafish retina can be regenerated with the help of myeloid cells to restore vision[164,165]. Another important advantage is that mice develop loose and nonnecrotic granulomas during M. tuberculosis infection, whereas zebrafish form lesions similar to human necrotic granulomas during M. marinum infection, making zebrafish well-suited for studying myeloid cells functions in granulomas[174]. Therefore, using zebrafish to model regeneration and disease provides a platform for investigating myeloid cell biology and developing related therapeutics, offering strong support for both basic research and clinical applications.
Advances using the zebrafish model can also be translated from bench to bedside. Introducing patient atypical gene fusion of PML-RARA to model acute promyelocytic leukemia (APL) in zebrafish enabled drug validation that confirmed the efficacy of classical APL drugs against this mutation, guiding clinical treatment and leading to complete remission in the patient[175]. In another notable application, Li et al. replicated an ARAF mutation from a patient in zebrafish, validated its pathogenesis effect, and screened a drug that successfully alleviated the patient’s symptoms[176]. Drug screening has also identified prostaglandin E2 as a regulator of HSC homeostasis during transplantation, which has advanced to clinical trials[177,178]. Beyond drug screening, screening different lineages of Shigella sonnei identified the most virulent lineage and related neutrophil responses[179], demonstrating the utility of zebrafish for pathogen evaluation. Taken together, the zebrafish advantages in genetic conservation, gene-editing, visualization, and drug/pathogen screening can aid clinical research related to myeloid cells.
Acknowledgments
Figure 1 and Figure 3 were respectively generated using the artificial intelligence tool GPT Image 2 and Nano Banana. The authors have reviewed, edited, and approved the final content and take full responsibility.
Authors contribution
Li G: Writing-original draft, visualization.
Lao J, Liu S: Investigation.
Deng J: Investigation, visualization.
Liang R: Investigation, visualization.
Zhang Y: Conceptualization, writing-review & editing.
Conflicts of interest
Yiyue Zhang is an Editorial Board Member of Myeloid Cells. The other author declares no conflicts of interest.
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Availability of data and materials
Not applicable.
Funding
This work was supported by the Guangdong Basic and Applied Basic Research Foundation (Grant Nos. 2024B1515040019 and 2026A1515012117).
Copyright
© The Author(s) 2026.
References
-
3. Grover A, Tcyganov EN, Gabrilovich DI. Myeloid cell reprogramming and immune suppression. Annu Rev Physiol. 2026;88:437-457.[DOI]
-
5. Patton EE, Zon LI, Langenau DM. Zebrafish disease models in drug discovery: From preclinical modelling to clinical trials. Nat Rev Drug Discov. 2021;20(8):611-628.[DOI]
-
7. Xue N, Liu X, Zhang D, Wu Y, Zhong Y, Wang J, et al. Improving adenine and dual base editors through introduction of TadA-8e and Rad51DBD. Nat Commun. 2023;14(1):1224.[DOI]
-
8. Zheng S, Liu Y, Xia X, Xiao J, Ma H, Yuan X, et al. Sequence context-agnostic TadA-derived cytosine base editors for genome-wide editing in zebrafish. Adv Sci. 2025;12(14):2411478.[DOI]
-
13. Kalvaitytė M, Balciunas D. Conditional mutagenesis strategies in zebrafish. Trends Genet. 2022;38(8):856-868.[DOI]
-
17. Kroll F, Powell GT, Ghosh M, Gestri G, Antinucci P, Hearn TJ, et al. A simple and effective F0 knockout method for rapid screening of behaviour and other complex phenotypes. ELife. 2021;10:e59683.[DOI]
-
18. Wu RS, Lam II, Clay H, Duong DN, Deo RC, Coughlin SR. A rapid method for directed gene knockout for screening in G0 zebrafish. Dev Cell. 2018;46(1):112-125.e4.[DOI]
-
23. Curado S, Anderson RM, Jungblut B, Mumm J, Schroeter E, Stainier DYR. Conditional targeted cell ablation in zebrafish: A new tool for regeneration studies. Dev Dyn. 2007;236(4):1025-1035.[DOI]
-
26. Li L, Yan B, Shi YQ, Zhang WQ, Wen ZL. Live imaging reveals differing roles of macrophages and neutrophils during zebrafish tail fin regeneration. J Biol Chem. 2012;287(30):25353-25360.[DOI]
-
27. Xu J, Zhu L, He S, Wu Y, Jin W, Yu T, et al. Temporal-spatial resolution fate mapping reveals distinct origins for embryonic and adult microglia in zebrafish. Dev Cell. 2015;34(6):632-641.[DOI]
-
29. Ellett F, Pase L, Hayman JW, Andrianopoulos A, Lieschke GJ. mpeg1 promoter transgenes direct macrophage-lineage expression in zebrafish. Blood. 2011;117(4):e49-e56.[DOI]
-
32. Ferrero G, Mahony CB, Dupuis E, Yvernogeau L, Di Ruggiero E, Miserocchi M, et al. Embryonic microglia derive from primitive macrophages and are replaced by cmyb-dependent definitive microglia in zebrafish. Cell Rep. 2018;24(1):130-141.[DOI]
-
35. Zhou Q, Zhao C, Yang Z, Qu R, Li Y, Fan Y, et al. Cross-organ single-cell transcriptome profiling reveals macrophage and dendritic cell heterogeneity in zebrafish. Cell Rep. 2023;42(7):112793.[DOI]
-
39. Jin H, Huang Z, Chi Y, Wu M, Zhou R, Zhao L, et al. C-Myb acts in parallel and cooperatively with Cebp1 to regulate neutrophil maturation in zebrafish. Blood. 2016;128(3):415-426.[DOI]
-
42. Wu Z, Hu Y, Xu S, Huang Z, Zhang W, Liu W. Expression and function of zebrafish carboxypeptidase A5 in neutrophils and mast cells. Fish Shellfish Immunol. 2026;173:111239.[DOI]
-
45. Herbert M, Goosmann C, Brinkmann V, Dimmler C, Cronan MR. Identification of a specific granular marker of zebrafish eosinophils enables development of new tools for their study. J Immunol. 2024;213(12):1893-1901.[DOI]
-
50. Farnsworth DR, Saunders LM, Miller AC. A single-cell transcriptome atlas for zebrafish development. Dev Biol. 2020;459(2):100-108.[DOI]
-
51. Lange M, Granados A, VijayKumar S, Bragantini J, Ancheta S, Kim YJ, et al. A multimodal zebrafish developmental atlas reveals the state-transition dynamics of late-vertebrate pluripotent axial progenitors. Cell. 2024;187(23):6742-6759.e17.[DOI]
-
55. Dobson JT, Seibert J, Teh EM, Da’as S, Fraser RB, Paw BH, et al. Carboxypeptidase A5 identifies a novel mast cell lineage in the zebrafish providing new insight into mast cell fate determination. Blood. 2008;112(7):2969-2972.[DOI]
-
57. Zhao C, Li Y, Tang J, Zhou Q, Lin X, Wen Z. Metaphocytes are IL-22BP-producing cells regulated by ETS transcription factor Spic and essential for zebrafish barrier immunity. Cell Rep. 2023;42(5):112483.[DOI]
-
58. de Jong JLO, Zon LI. Use of the zebrafish system to study primitive and definitive hematopoiesis. Annu Rev Genet. 2005;39:481-501.[DOI]
-
62. Dai Y, Zhu L, Huang Z, Zhou M, Jin W, Liu W, et al. Cebpα is essential for the embryonic myeloid progenitor and neutrophil maintenance in zebrafish. J Genet Genom. 2016;43(10):593-600.[DOI]
-
68. He Y, Li Y, Shao Y, Huang X, He J, Liu S, et al. Regulation of an lncRNAirf8by the Ikzf1/Myb complex drives neutrophil development. Proc Natl Acad Sci U S A. 2025;122(41):e2502741122.[DOI]
-
74. Wang Y, Yang R, Wang P, Liu X, Yuan H, Huang H, et al. Tead1a initiates transcriptional priming through the TEAD1a/YAP-Notch1-Spi1/cebpα axis to promote neutrophil fate. Adv Sci. 2025;12(41):e05441.[DOI]
-
75. Yan L, Tan S, Wang H, Yuan H, Liu X, Chen Y, et al. Znf687 recruits Brd4-Smrt complex to regulate gfi1aa during neutrophil development. Leukemia. 2024;38(4):851-864.[DOI]
-
76. Hall CJ, Flores MV, Oehlers SH, Sanderson LE, Lam EY, Crosier KE, et al. Infection-responsive expansion of the hematopoietic stem and progenitor cell compartment in zebrafish is dependent upon inducible nitric oxide. Cell Stem Cell. 2012;10(2):198-209.[DOI]
-
77. Hogan BM, Layton JE, Pyati UJ, Nutt SL, Hayman JW, Varma S, et al. Specification of the primitive myeloid precursor pool requires signaling through Alk8 in zebrafish. Curr Biol. 2006;16(5):506-511.[DOI]
-
78. Arroyo AB, Tyrkalska SD, Bastida-Martínez E, Monera-Girona AJ, Cantón-Sandoval J, Bernal-Carrión M, et al. Peds1 deficiency in zebrafish results in myeloid cell apoptosis and exacerbated inflammation. Cell Death Discov. 2024;10:388.[DOI]
-
80. Campbell CA, Fursova O, Cheng X, Snella E, McCune A, Li L, et al. A zebrafish model of granulin deficiency reveals essential roles in myeloid cell differentiation. Blood Adv. 2021;5(3):796-811.[DOI]
-
84. Lazarov T, Juarez-Carreño S, Cox N, Geissmann F. Physiology and diseases of tissue-resident macrophages. Nature. 2023;618(7966):698-707.[DOI]
-
85. Xu J, Wang T, Wu Y, Jin W, Wen Z. Microglia colonization of developing zebrafish midbrain is promoted by apoptotic neuron and lysophosphatidylcholine. Dev Cell. 2016;38(2):214-222.[DOI]
-
89. Liu W, Di Q, Li K, Li J, Ma N, Huang Z, et al. The synergistic role of Pu.1 and Fms in zebrafish osteoclast-reducing osteopetrosis and possible therapeutic strategies. J Genet Genom. 2020;47(9):535-546.[DOI]
-
100. Nasri M, Dannenmann B, Doll L, Findik B, Bernhard F, Kandabarau S, et al. Flavopiridol restores granulopoiesis in experimental models of severe congenital neutropenia. Mol Ther. 2025;33(6):2851-2871.[DOI]
-
101. Borbaran Bravo N, Deordieva E, Doll L, ElGamacy M, Dannenmann B, Azevedo J, et al. A new severe congenital neutropenia syndrome associated with autosomal recessive COPZ1 mutations. Blood. 2025;145(20):2317-2335.[DOI]
-
102. Konantz M, Schürch C, Hanns P, Müller JS, Sauteur L, Lengerke C. Modeling hematopoietic disorders in zebrafish. Dis Models Mech. 2019;12(9):dmm040360.[DOI]
-
103. Rajan V, Collett K, Woodside R, Prykhozhij SV, Moksa M, Carles A, et al. Stress hematopoiesis induces a proliferative advantage in TET2 deficiency. Leukemia. 2022;36(3):809-820.[DOI]
-
105. Tu L, He F, Liew JM, Sin CF, Zheng L, Tsui SP, et al. asxl1 C-terminal truncation and SRSF2 mutation drive leukemogenesis via immune reprogramming. Leukemia. 2025;39(12):3047-3051.[DOI]
-
108. Xu SE, Hong J, Dongye M, Lin J, Xue R, Huang Z, et al. The synergistic effect of c-Myb hyperactivation and Pu.1 deficiency induces Pelger–Huët anomaly and promotes sAML. Proc Natl Acad Sci U S A. 2025;122(9):e2416121122.[DOI]
-
109. Travnickova J, Tran Chau V, Julien E, Mateos-Langerak J, Gonzalez C, Lelièvre E, et al. Primitive macrophages control HSPC mobilization and definitive haematopoiesis. Nat Commun. 2015;6:6227.[DOI]
-
114. Yang RM, Song SY, Wu FY, Yang RF, Shen YT, Tu PH, et al. Myeloid cells interact with a subset of thyrocytes to promote their migration and follicle formation through NF-κB. Nat Commun. 2023;14:8082.[DOI]
-
115. Hu YX, You HM, Bai MR, Yue WH, Li FF, Hu BW, et al. Macrophage P2Y12 regulates iron transport and its inhibition protects against atherosclerosis. J Adv Res. 2025;76:585-603.[DOI]
-
117. Bravo P, Liu Y, Draper BW, Marlow FL. Macrophage activation drives ovarian failure and masculinization in zebrafish. Sci Adv. 2023;9(47):eadg7488.[DOI]
-
123. Gomes MC, Brokatzky D, Mostowy S. Shigella-trained pro-inflammatory macrophages protect zebrafish from secondary infection. Cell Rep. 2025;44(5):115601.[DOI]
-
127. Luo G, Wen Y, Wang M, Wang H, Li D, Liao M, et al. A granulin-positive macrophage subtype in mycobacterial granulomas alleviates tissue damage by limiting excessive inflammation. Proc Natl Acad Sci U S A. 2025;122(31):e2413946122.[DOI]
-
131. Clay H, Davis JM, Beery D, Huttenlocher A, Lyons SE, Ramakrishnan L. Dichotomous role of the macrophage in early mycobacterium marinum infection of the zebrafish. Cell Host Microbe. 2007;2(1):29-39.[DOI]
-
132. Viswanathan G, Hughes EJ, Gan M, Xet-Mull AM, Lowy JP, Pyle CJ, et al. Granuloma dual RNA-seq reveals composite transcriptional programs driven by neutrophils and necrosis within tuberculous granulomas. Sci Adv. 2026;12(4):eadw4619.[DOI]
-
133. Hu CB, Wang J, Hong Y, Li H, Fan DD, Lin AF, et al. Single-cell transcriptome profiling reveals diverse immune cell populations and their responses to viral infection in the spleen of zebrafish. FASEB J. 2023;37(6):e22951.[DOI]
-
134. Roux E, Willms RJ, Van Dycke J, Cortes Calabuig Á, Van Espen L, Schoofs G, et al. Transcriptional profiling of zebrafish intestines identifies macrophages as host cells for human norovirus infection. Gut Microbes. 2024;16:2431167.[DOI]
-
147. Xie Y, Meijer AH, Schaaf MJM. Modeling inflammation in zebrafish for the development of anti-inflammatory drugs. Front Cell Dev Biol. 2021;8:620984.[DOI]
-
148. Brugman S, Liu K, Lindenbergh–Kortleve D, Samsom JN, Furuta GT, Renshaw SA, et al. Oxazolone-induced enterocolitis in zebrafish depends on the composition of the intestinal microbiota. Gastroenterology. 2009;137(5):1757-1767.e1.[DOI]
-
149. Sanmarco LM, Chao CC, Wang YC, Kenison JE, Li Z, Rone JM, et al. Identification of environmental factors that promote intestinal inflammation. Nature. 2022;611(7937):801-809.[DOI]
-
154. Marques IJ, Lupi E, Mercader N. Model systems for regeneration: Zebrafish. Development. 2019;146(18):dev167692.[DOI]
-
155. Kawakami A, Fukazawa T, Takeda H. Early fin primordia of zebrafish larvae regenerate by a similar growth control mechanism with adult regeneration. Dev Dyn. 2004;231(4):693-699.[DOI]
-
158. Zareba J, Cattaneo EF, Villani A, Othman A, Streb S, Peri F. NPC1 links cholesterol trafficking to microglial morphology via the gastrosome. Nat Commun. 2024;15:8638.[DOI]
-
160. Zhao C, Lin G, Cheng K, Hassen S, Pan H, Wen Z. Swimming through dendritic cell biology: Insights from fish models. Immunol Rev. 2025;336:e70067.[DOI]
-
166. Poss KD, Wilson LG, Keating MT. Heart regeneration in zebrafish. Science. 2002;298(5601):2188-2190.[DOI]
-
170. Simões FC, Cahill TJ, Kenyon A, Gavriouchkina D, Vieira JM, Sun X, et al. Macrophages directly contribute collagen to scar formation during zebrafish heart regeneration and mouse heart repair. Nat Commun. 2020;11:600.[DOI]
-
173. Sato S, Kefalov VJ. Characterization of zebrafish rod and cone photoresponses. Sci Rep. 2025;15:13413.[DOI]
-
175. Ye Y, Zhao Z, Mo W, Liu W, Wu L, Li J, et al. Zebrafish modeling of atypical PML-RARA isoform from acute promyelocytic leukemia patient and its implications for clinical treatment. Ann Hematol. 2025;104(1):171-181.[DOI]
-
176. Li D, March ME, Gutierrez-Uzquiza A, Kao C, Seiler C, Pinto E, et al. ARAF recurrent mutation causes central conducting lymphatic anomaly treatable with a MEK inhibitor. Nat Med. 2019;25(7):1116-1122.[DOI]
Copyright
© The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
Publisher’s Note
Science Exploration remains a neutral stance on jurisdictional claims in published
maps
and institutional affiliations. The views expressed in this article are solely those
of
the author(s) and do not reflect the opinions of the Editors or the publisher.
Share And Cite



Science Exploration Style
Li G, Lao J, Deng J, Liu S, Liang R, Zhang Y. Zebrafish: A window into myeloid cell development, disease, and defense. Myeloid Cells. 2026;1:202505. https://doi.org/10.70401/mc.2026.0002
Tips
Copy completed.
Submit a Manuscript
Author Instructions
Cite this Article
Article Metrics
0
View
0
Download
Cited
Article Updates

- Abstract
- Keywords
- 1. Introduction
- 2. Why Zebrafish?
- 3. Zebrafish Myeloid Cells
- 4. Zebrafish Myelopoiesis
- 5. Zebrafish Myeloid Cell Function
- 6. Conclusion
- Acknowledgments
- Authors contribution
- Conflicts of interest
- Ethical approval
- Consent to participate
- Consent for publication
- Availability of data and materials
- Funding
- References
- Copyright
Science Exploration Style
Li G, Lao J, Deng J, Liu S, Liang R, Zhang Y. Zebrafish: A window into myeloid cell development, disease, and defense. Myeloid Cells. 2026;1:202505. https://doi.org/10.70401/mc.2026.0002
copy
Share Link
copy