Neutrophils in cancer: Insights from intravital imaging
Sapna Devi
*

*Correspondence to:
Sapna Devi, Department of Microbiology and Immunology, The University of Melbourne, The Peter Doherty Institute for Infection and Immunity, Melbourne, VIC 3000, Australia.
E-mail: sapna.devi@unimelb.edu.au
Myeloid Cells. 2026;1:202607. 10.70401/mc.2026.0007
Received: April 05, 2026Accepted: June 29, 2026Published: June 30, 2026
Abstract
Multiple clinical and translational studies have shown that infiltration of tumour-associated neutrophils is associated with advanced disease stages, increased metastatic spread, therapeutic resistance, and poorer overall survival across various cancer types. Beyond their traditional role as first responders to tissue damage and pathogen infection, neutrophils within the tumour microenvironment (TME) display remarkable functional plasticity, adopting phenotypes that can promote angiogenesis, extracellular matrix remodelling, immune suppression, and tumour cell invasion. Importantly, it is now recognised that the prognostic impact of neutrophils is highly context-dependent, shaped by tumour type, stage, and the evolving inflammatory milieu. Intravital imaging studies have revealed dynamic neutrophil behaviours, including distinct migratory patterns between intra-tumoural and peri-tumoural regions, interactions with tumour and immune cells, and contributions to processes such as metastasis and immune suppression. However, there is a lack of understanding of how transcriptionally defined neutrophil subsets translate into specific functional and behavioural states in the TME in vivo. This mini-review spotlights intravital imaging approaches that illuminate neutrophil dynamics in tumours. We also discuss how extrinsic regulators, including cancer-associated fibroblasts and neural inputs, direct neutrophil dynamics, further contributing to TME complexity.
Keywords
Neutrophils, intravital imaging, tumour microenvironment, cancer
1. Introduction
Intravital imaging serves as a powerful approach for achieving visual representation and quantitative analysis at both the cellular and subcellular levels in vivo. This technique allows longitudinal tracking of individual cells within intact organs of living animals and has helped identify a number of previously unappreciated mechanisms operating in the steady-state[1-3] and disease settings[4-7], thereby driving fundamental advances across fields including immunology, cancer biology, and developmental biology.
While many contemporary methodologies such as flow cytometry, multiplex confocal imaging, and single cell transcriptomics of tissues and cells have greatly progressed clinical and biomedical research, these only provide a static snapshot of the occurring molecular and/or cellular changes. With the exception of multi-parameter labelling methods combined with tissue imaging[8,9], many techniques overlook the spatial and temporal distributions of cells and their interactions during disease development. In contrast, intravital imaging enables real-time visualization of cell trafficking, cell–cell interactions, and functional responses within intact tissues. By preserving physiological blood flow, neural inputs, and stromal architecture, intravital imaging has revealed unexpected leukocyte behaviours, including rapid neutrophil swarming[10,11], prolonged tissue residency[12,13], and dynamic interactions with tumour, endothelial, and immune cells[14-18] in a variety of tissues. This mini review summarises the latest advances in understanding the role of neutrophils in cancer, with a specific focus on using live intravital imaging to elucidate their behaviour and functionality during an anti-tumour response. An improved insight into the roles of neutrophils within tumours will reveal new mechanisms controlling their recruitment and function, informing the development of neutrophil-targeted immunotherapeutic strategies.
2. Intravital Imaging Modalities
Technological breakthroughs in imaging over the past few decades have revolutionised in vivo visualization, enabling unprecedented insight into dynamic biological processes. High-resolution methods for intravital imaging include confocal laser-scanning microscopy, spinning-disc confocal microscopy, and multiphoton microscopy. These modalities differ in illumination strategy, tissue penetration, spatial resolution, imaging area, and the degree of photobleaching and phototoxicity. The imaging modality chosen should depend on the biological constraints. Confocal microscopy uses a pinhole placed before the detector to block out-of-focus light, enhancing image contrast and spatial resolution[19]. Additionally, spinning disk confocal microscopy increases imaging speed by using multiple pinholes for simultaneous multi-point scanning, making it well suited for capturing dynamic cellular processes in real time[2]. However, both of these methods use single-photon excitation of fluorophores with excitation wavelengths (typically 400-800 nm) that limit deep imaging into tissues. In contrast, multiphoton microscopy relies on the near-simultaneous absorption of low-energy pulsed infrared light (typically 800-1,200 nm) to excite fluorophores, which allows for imaging deeper into tissue while maintaining high spatial resolution[19]. Both approaches produce high-resolution images, but multiphoton techniques are particularly advantageous for intravital imaging of thick or scattering samples. This is due to the longer excitation wavelengths that penetrate deeper into tissues and are less scattered than shorter wavelengths. In addition, fluorescence excitation occurs only at the focal point, where two (or more) photons are simultaneously absorbed. This spatially restricted excitation minimizes out-of-focus signal, reduces photodamage to surrounding tissue, and improves imaging depth and resolution in intact, living samples. These improvements in intravital imaging have opened a window into the dynamic processes unfolding within living tissues.
In practice, the performance of intravital imaging is determined not only by optical modality but also by the biological preparation, imaging window, fluorophore brightness, and the balance between temporal resolution and tissue preservation. Depending on the tissue and experimental setup, single imaging sessions can range from tens of minutes to several hours, while chronic imaging window approaches may permit repeated visualization of the same tumour region over multiple days or longer[1,3,20,21]. Typical fields of view range from hundreds of microns to low millimetres, depending on the objective and acquisition settings, and imaging depth is generally more limited for confocal and spinning-disc approaches than for multiphoton microscopy, which is better suited for imaging deeper within scattering tumour tissues. However, attempts to extend imaging duration or depth often require higher laser power or longer exposure, increasing the risk of photobleaching, phototoxicity, local thermal damage, and artifactual sterile inflammation. These issues are particularly important when studying neutrophils, as they are highly sensitive to tissue perturbation and may rapidly respond to laser-induced damage, thereby confounding interpretation of their behaviour within the tumour microenvironment.
To study immune cells, including neutrophils, and their surrounding tissue microenvironment, intravital microscopy relies on in vivo cellular and molecular labelling strategies. These include fluorescently conjugated antibodies, genetically encoded fluorescent proteins, specialised photoconvertible reporters such as Kaede[22], and genetically engineered reporter mice expressing cell-specific fluorescent markers. Together, these approaches enable precise identification of distinct cell populations, tracking of their migration, and real-time analysis of their dynamic behaviours within intact tissues. Such strategies allow us not only to distinguish immune and stromal subsets, but also to interrogate how cells interact, respond to environmental cues, and remodel their surroundings over time. In addition, intravital imaging can be coupled with genetically encoded biosensors that report cellular activation and signalling states in vivo, thereby extending analysis beyond cell localization and motility alone. Examples include calcium-sensitive indicators such as GCaMP, enabling visualization of rapid intracellular calcium flux[21,23,24] and transcriptional reporters such as nuclear factor of activated T cells (NFAT)-based systems[25,26] which provide readouts of downstream activation over longer timescales. Successful application of these approaches, particularly during high-resolution imaging of rapidly migrating cells such as neutrophils, also depends on effective tissue stabilization to minimise motion artefacts caused by respiration, heartbeat, and tissue drift. This is commonly achieved using imaging windows, tissue restraints, or suction-based platforms depending on the tissue and experimental setting[20]. In addition to these commonly used modalities, other optical approaches can provide complementary information in selected settings. For example, fluorescence lifetime imaging microscopy may offer insight into metabolic or signalling states, whereas total internal reflection fluorescence microscopy is more suited to near-surface events and is therefore less broadly applicable to intravital imaging of intact solid tumours. Beyond fluorescent labelling, advanced imaging modalities such as multiphoton microscopy provide additional structural information through nonlinear optical processes, including second-harmonic generation (SHG) and third-harmonic generation (THG)[1,19]. These label-free signals arise from intrinsic tissue properties, enabling visualization of tissue structures without exogenous dyes. SHG highlights highly ordered structures such as collagen fibres, revealing extracellular matrix (ECM) organization, while THG detects interfaces and lipid-rich structures, offering insight into tissue architecture and cellular morphology. When combined with fluorescent detection of labelled cells, these complementary signals enable deep tissue, high-resolution three-dimensional imaging, providing a comprehensive and dynamic view of cellular behaviour within its structural microenvironment. As contemporary research increasingly relies on diverse and high-dimensional techniques, intravital imaging using the preferred choice of imaging modality plays a critical complementary role by providing in vivo context in health and disease.
3. Neutrophils: Good or Bad in Cancer?
Neutrophils are the most abundant circulating leukocytes and represent a first line of defence against infection and tissue injury. Traditionally viewed as short-lived, terminally differentiated effector cells, neutrophils are now recognised as highly dynamic and functionally diverse, with roles extending beyond antimicrobial defence to include regulation of inflammation, tissue remodelling, and cancer progression[27-30]. A high intratumoural frequency of neutrophils has emerged as a strong predictor of poor clinical outcome across the majority of solid malignancies[31-34]. Neutrophil polarization is likely a key determinant of the functional roles they assume within the tumour microenvironment (TME). Rather than representing a uniform population, neutrophils exhibit context-dependent phenotypic and functional diversity shaped by local cytokines, chemokines, metabolic cues, and cellular interactions. This polarization may dictate whether they adopt anti-tumour effector functions or, conversely, promote tumour growth, immune suppression, angiogenesis, and metastasis.
In 2009, Fridlender et al., proposed a functional dichotomy of tumour-associated neutrophils distinguishing anti-tumourigenic (N1) from pro-tumourigenic (N2) phenotypes. This nomenclature was inspired by the polarization paradigm originally established for macrophages as pro-inflammatory (M1) and anti-inflammatory (M2). The study demonstrated that transforming growth factor-β (TGF-β), a cytokine frequently overexpressed in tumours and known for its immunosuppressive properties, could skew neutrophils toward a pro-tumour (N2) state[35]. In mouse models, depletion of neutrophils resulted in a reduction in tumour growth, suggesting their potential contribution to tumour progression. Notably, blockade of TGF-β signalling led to the accumulation of neutrophils with enhanced cytotoxicity and anti-tumour activity, consistent with an N1-like phenotype. These findings provided early evidence that neutrophil function in cancer is not fixed but can be reprogrammed by signals within the TME. Despite their reported functional differences, no definitive or universally accepted surface markers have been identified that reliably distinguish N1 from N2 neutrophils. This lack of clear phenotypic delineation suggests that these subpopulations may not represent stable, discrete lineages, but rather context-dependent activation states along a dynamic spectrum of neutrophil plasticity. Consequently, the N1/N2 framework, like the current view of macrophage subsets[36], is best viewed as a functional model reflecting differential activation and environmental conditioning within the TME, rather than as strictly separable cellular subsets.
Around the same time, myeloid-derived suppressor cells (MDSCs) were formally described as a heterogenous population of pathologically activated myeloid cells enriched in cancer patients and characterised by potent immunosuppressive activity[37,38]. These cells were shown to inhibit T cell proliferation and effector function through mechanisms including arginase-1 activity, reactive oxygen species production, nitric oxide generation, and cytokine-mediated suppression. Importantly, granulocytic, or polymorphonuclear MDSCs (PMN-MDSCs) share phenotypic and functional similarities with neutrophils, further blurring the distinction between classical neutrophils and tumour-promoting myeloid subsets. This conceptual shift expanded the understanding of neutrophil biology in cancer, highlighting their capacity not only to mediate acute inflammation but also to contribute to immune evasion and tumour progression within the TME. A persistent challenge in the field is the distinction between classical neutrophils, tumour-associated neutrophil states, and PMN-MDSCs. These populations share substantial phenotypic and functional overlap, particularly in mouse models, where surface markers and morphology often do not permit clear separation. For this reason, terms such as "subset," "state," and "phenotype" should be used carefully. In this review, we refer to transcriptionally or experimentally separable groups as neutrophil populations or subsets where appropriate, while using "state" to describe dynamic functional programs shaped by the TME. This distinction is important because many tumour-associated neutrophil features, including immunosuppression, prolonged survival, and pro-angiogenic activity, may reflect environmentally induced states rather than discrete lineages.
Single-cell transcriptomic approaches have provided unprecedented resolution into neutrophil heterogeneity, revealing diverse transcriptional clusters within tumours and other inflammatory settings[28,30,39-44]. For example, in a murine model of 4T1 breast cancer lung metastasis, neutrophils derived from the bone marrow or circulation exhibited little to no immunosuppressive capacity[44]. In contrast, neutrophils that had infiltrated metastatic lung lesions displayed potent suppressive activity against both T cells and natural killer (NK) cells. Transcriptional profiling revealed three subpopulations of lung-resident neutrophils. Notably, in tumour-free mice only two subsets were detectable, whereas mice bearing 4T1 tumours demonstrated a marked expansion of a third, tumour-associated population. Furthermore, all lung-infiltrating neutrophil clusters showed elevated expression of genes linked to immunosuppression, including Ptgs2, IL1b, IL10, Arg1, Arg2, Nos2, CD274, and Trem1 when compared with their blood or bone marrow counterparts, underscoring the profound influence of the TME on neutrophil functional reprogramming. In a separate study, Ng and colleagues showed that both mature and immature neutrophils derived from the bone marrow are capable of infiltrating tumours in a mouse model of pancreatic ductal adenocarcinoma. Once within the TME, these populations undergo distinct epigenetic and transcriptional reprogramming, giving rise to two intermediate states termed T1 and T2. As tumour progression continues, further environmental conditioning drives both subsets toward a convergent, terminally differentiated state referred to as T3[28]. In mice, T3 neutrophils are marked by expression of dcTRAIL-R1, exhibit a markedly prolonged lifespan exceeding five days, and display pro-angiogenic as well as pro-tumoural activities. Importantly, several defining features of the T3 population, including enhanced survival and elevated vascular endothelial growth factor (VEGF) production, mirror properties previously associated with N2-polarised neutrophils, suggesting potential overlap between these conceptual frameworks of neutrophil functional states. These findings suggest that neutrophils comprise multiple molecularly distinct populations, each potentially endowed with specialised functional programs.
Rather than existing as a uniform cell type, neutrophils appear to span a spectrum of states associated with discrete biological roles, ranging from inflammatory effector functions to immunoregulatory and tissue-remodelling activities. However, whether this molecular heterogeneity translates into functionally and behaviourally distinct subsets within the TME remains unclear. Determining how these transcriptionally defined states manifest in terms of migration dynamics, cellular interactions, spatial positioning, and lifespan in vivo requires direct visualization approaches. In this context, intravital imaging offers a powerful platform to bridge molecular identity with real-time behaviour, enabling investigation of how discrete neutrophil states operate within the living TME.
4. Imaging the Tumour Microenvironment
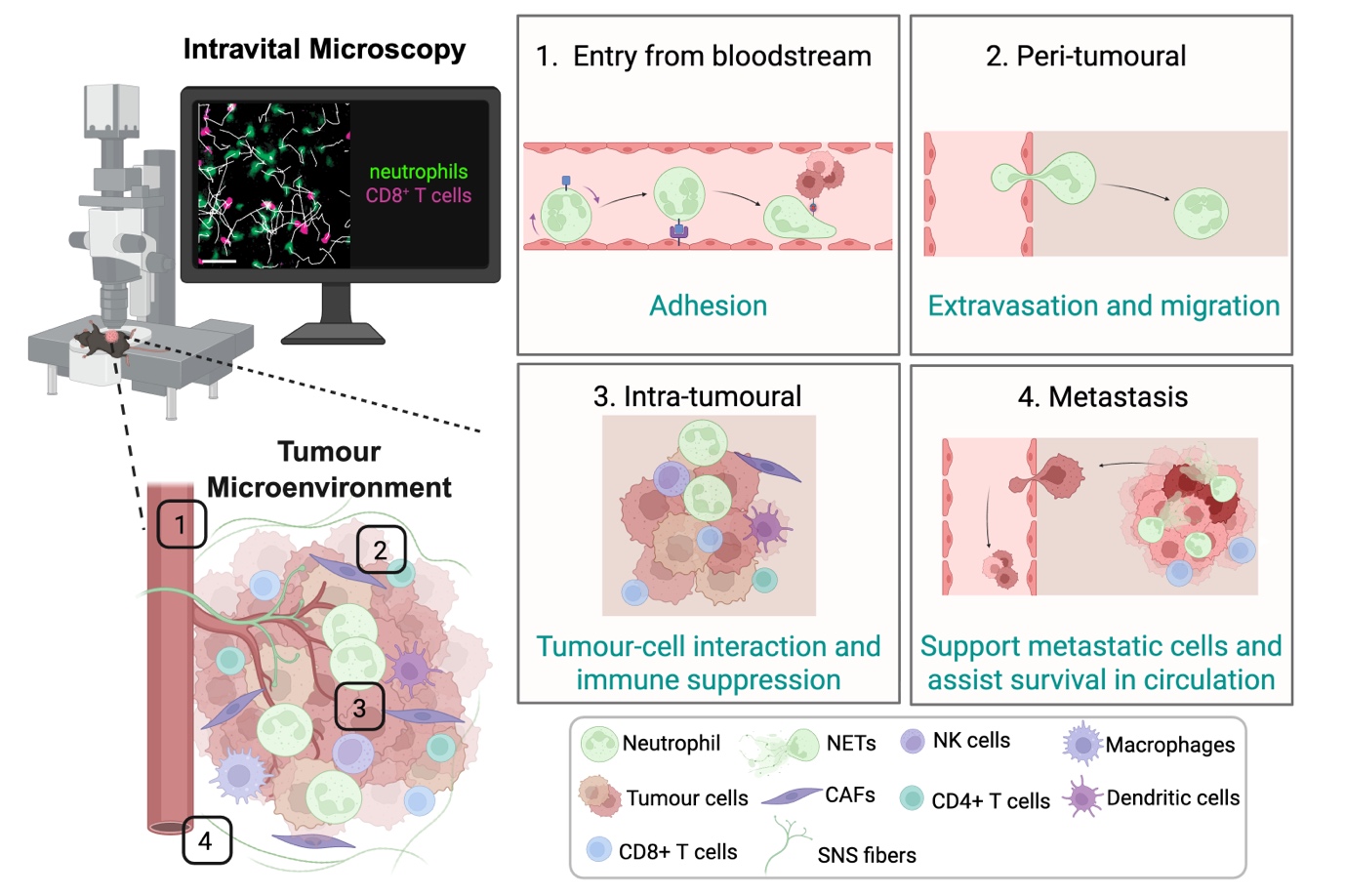
In most solid tumours, the TME is a highly dynamic and complex ecosystem composed of malignant cells, stromal elements, vasculature, ECM, and a diverse milieu of immune populations. Within this intricate network, continuous cellular crosstalk determines whether anti-tumour immunity is successfully mounted or whether tumour progression and immune evasion prevail. There are many reasons that contribute to this balance, including accumulating evidence indicating that disease outcome is not dictated solely by tumour-intrinsic factors, but rather by the evolving interactions between cancer cells and their surrounding microenvironment. Effective tumour immunosurveillance requires the coordinated migration, infiltration, and sustained function of effector immune cells within the tumour bed. Intravital imaging has been instrumental in revealing how these encounters are either facilitated or constrained in real time (Figure 1). For example, in a melanoma model, patrolling epidermal resident memory T cells help sustain cancer–immune equilibrium[45]. Intravital imaging studies in various cancer models have revealed that cytotoxic T cell (CTL) migration exhibits distinct behaviours across regions of the TME, from the periphery to the tumour core, highlighting the dynamic factors that regulate T cell motility[14,46-49]. Stable contacts between cytotoxic T lymphocytes (CTLs) and tumour cells are essential for effective killing[14,15], whereas physical barriers such as dense collagen networks can restrict immune cell access and limit therapeutic efficacy[50,51].
{kind=link}
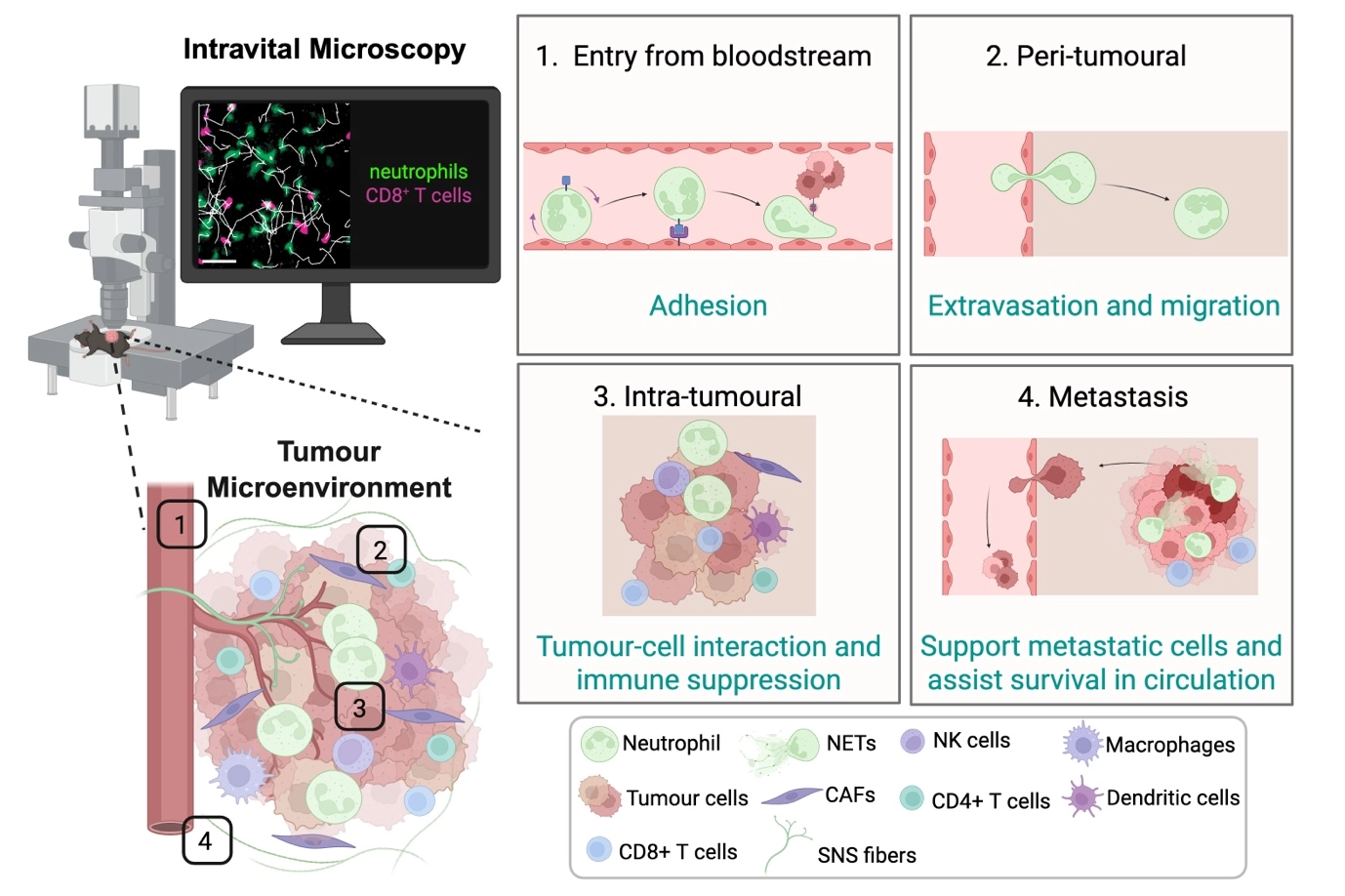
Figure 1. Intravital imaging of different locations in the TME reveals dynamic cell behaviours. Cells are recruited from the bloodstream and initially undergo adhesion to the endothelium (1), followed by extravasation and migration into peri-tumoural regions (2); Within the tumour, immune cells exhibit dynamic intra-tumoural migration and interactions with tumour and/or immune cells, including CD8⁺ T cells, contributing to immune modulation and suppression (3); Beyond the primary tumour, some immune cell types can facilitate metastatic dissemination, supporting tumour cell survival in circulation and promoting colonization at distant sites (4); Together, these observations illustrate the spatial and temporal complexity of immune cell function in cancer, as revealed by live imaging approaches. Created in BioRender. Devi, S. (2026) https://BioRender.com/kuz17lw. TME: tumour microenvironment; NETs: neutrophil extracellular traps; NK: natural killer; CAFs: cancer-associated fibroblasts; SNS: sympathetic nervous system.
Similarly, intravital approaches have uncovered how myeloid populations dynamically interact within the TME, that may influence cellular positioning and shape anti-tumour immunity. In a model of spontaneous breast cancer, rare activating dendritic cells were discovered at the tumour margin that could compete with tumour-associated macrophages for T cell interactions, favouring tumour elimination[52]. In a separate study, it was demonstrated that macrophages of distinct origins occupy spatially segregated niches within tumours, where their behaviours can influence local immune interactions[53]. Together, these insights underscore how spatial organization and temporal cellular interactions within the TME critically influence the balance between immune control and tumour escape.
It has long been recognised that within the TME, neutrophils can exert cytotoxic effects through the production of reactive oxygen species (ROS) and nitric oxide (NO), as well as through the secretion of pro-inflammatory cytokines such as tumour necrosis factor-alpha (TNF-α)[29]. These mechanisms contribute to classical anti-tumour immune responses. However, more recent studies have revealed a far more nuanced role for neutrophils in cancer. Rather than acting solely as tumouricidal effector cells, neutrophils can also adopt pro-tumourigenic functions that promote tumour growth, angiogenesis, immune suppression, and metastatic dissemination[29,54]. This emerging duality highlights the functional plasticity of neutrophils within the TME. Here, we discuss the dynamic and context-dependent roles of neutrophils in cancer progression, with particular emphasis on insights gained through intravital microscopy, which has enabled real-time visualization of neutrophil behaviour within living tumours.
5. Neutrophil Dynamics in Cancer Revealed by Imaging
In solid tumours and inflamed tissues, neutrophils exhibit remarkable plasticity, rapidly adapting their migratory behaviour, lifespan, and effector functions in response to local microenvironmental cues. However, many of these behaviours are transient and spatially restricted, making them difficult to capture using conventional ex vivo analyses or static imaging of fixed tissues. A comprehensive understanding of cellular function requires approaches that capture not only molecular identity but also behaviour, context, and timing within living tissues. Using intravital imaging, neutrophils have been visualised in a number of tissues such as skin[10,11,55], bone marrow[56,57], lung[56,58], liver[59,60], and kidney[17,61]. To enable longitudinal tracking of neutrophils in vivo, several strategies are available, including adoptive transfer of pre-labelled neutrophils, systemic administration of fluorescently conjugated antibodies such as anti-Ly6G, and the use of genetically engineered reporter mice expressing fluorescent proteins in myeloid cells. Each approach has distinct strengths and limitations. For example, LysM-GFP mice enable robust visualization of myeloid cells, but fluorescence is not restricted to neutrophils and may also label monocytes and macrophages[62]. Anti-Ly6G-based labelling can provide selective visualization of neutrophils, although antibody dose and experimental design should be carefully considered, as perturbations of neutrophil behaviour or survival may confound interpretation. An important advance in the field has been the development of Catchup-based neutrophil reporter models, which provide greater specificity for neutrophil imaging in vivo[63]. For intravital imaging applications requiring brighter signals, this system can be combined with suitable Rosa26-based reporter alleles to improve fluorescence intensity and facilitate high-resolution visualization in vivo. More recently, a CD101 reporter mouse coupled to tdTomato fluorescence has emerged as an additional approach to identify and track mature neutrophils in vivo[64]. Applied in the context of intravital imaging, such a model may provide further insight into how neutrophil maturation state influences tissue positioning, migratory dynamics, and functional behaviour within the TME. These models are particularly well suited for long-term intravital imaging, as they permit continuous, non-invasive monitoring of neutrophil migration, accumulation, and cellular interactions within intact tissues over extended periods.
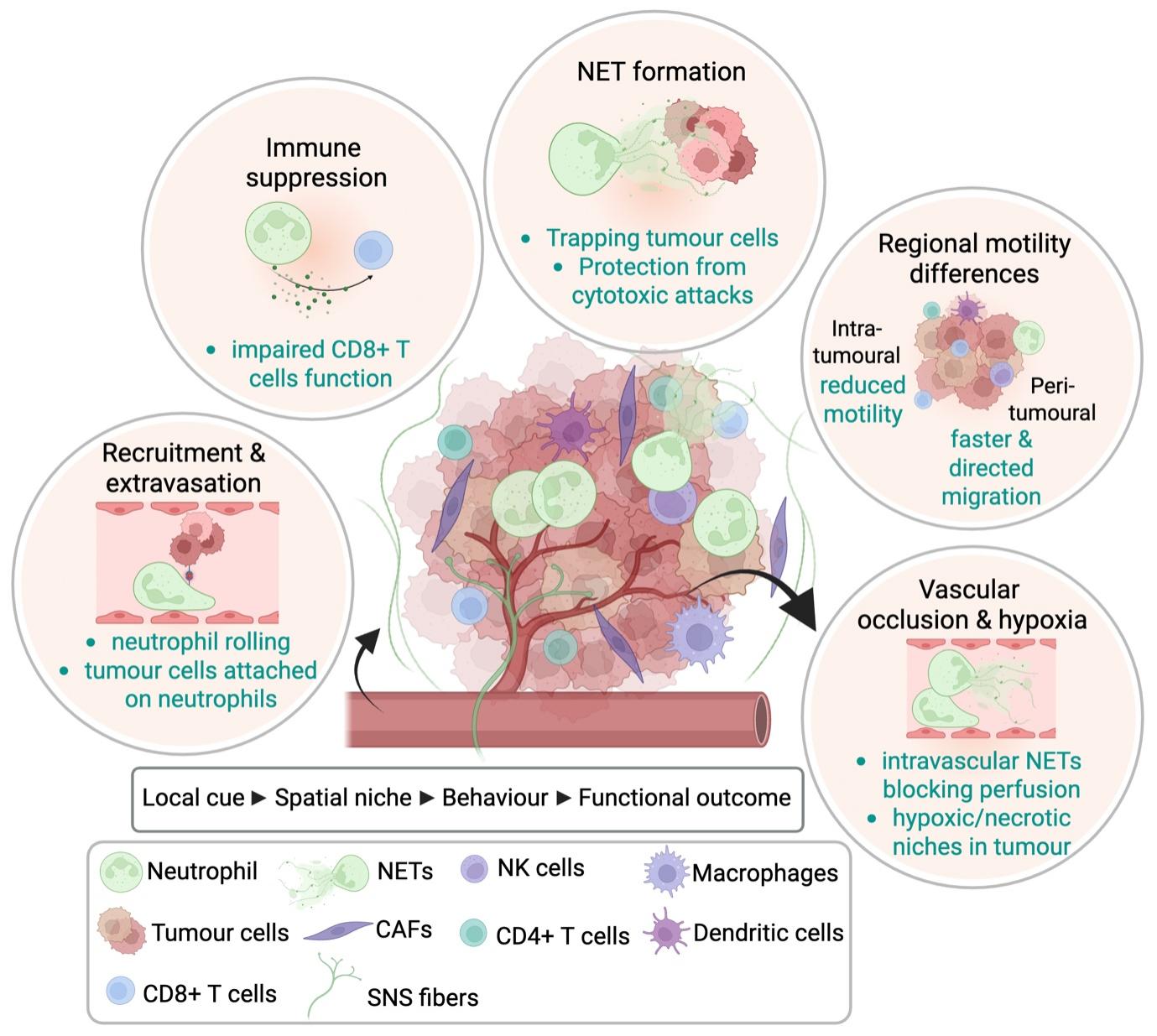
Early applications of intravital microscopy in tumour models focused predominantly on T cell dynamics[4,5,14]. Subsequent studies have extended this approach to neutrophils, revealing a range of dynamic and context-dependent behaviours within tumours that are not readily captured by static approaches (Table 1). These include recruitment from the vasculature, extravasation into peri-tumoural tissue, differential motility across peri- and intra-tumoural regions, direct interactions with tumour cells and other immune populations, participation in metastatic seeding, and the formation of neutrophil extracellular traps within primary and metastatic sites (Figure 2). A major strength of intravital imaging is its ability to directly visualize neutrophil recruitment from the circulation, including adhesion to the endothelium, extravasation, and early tissue entry within peri-tumoural regions. Prior work in hepatic cancer models using Lewis lung carcinoma cells demonstrated that neutrophils labelled via systemic antibody administration interact with circulating cancer cells within the liver sinusoids[59,60]. Intravital imaging of the liver microcirculation showed cancer cells arrest from the bloodstream directly over neutrophils attached to the sinusoidal endothelium within minutes of cancer cell injection, particularly during the first hour after dissemination[60]. These findings suggest that neutrophils can facilitate the adhesion of cancer cells within the TME and may support the initiation of metastasis. In another study, intravital imaging of established subcutaneous tumours generated using CT-26 cells enabled the visualization of neutrophils within the TME[65]. Importantly, this work was among the first to demonstrate that intravital imaging could be used not only to observe immune cell–tumour interactions in vivo, but also to quantitatively assess dynamic neutrophil behaviours. Parameters such as cell velocity, migration distance, and motility patterns were measured, providing quantitative insight into neutrophil dynamics within the TME.
{kind=link}
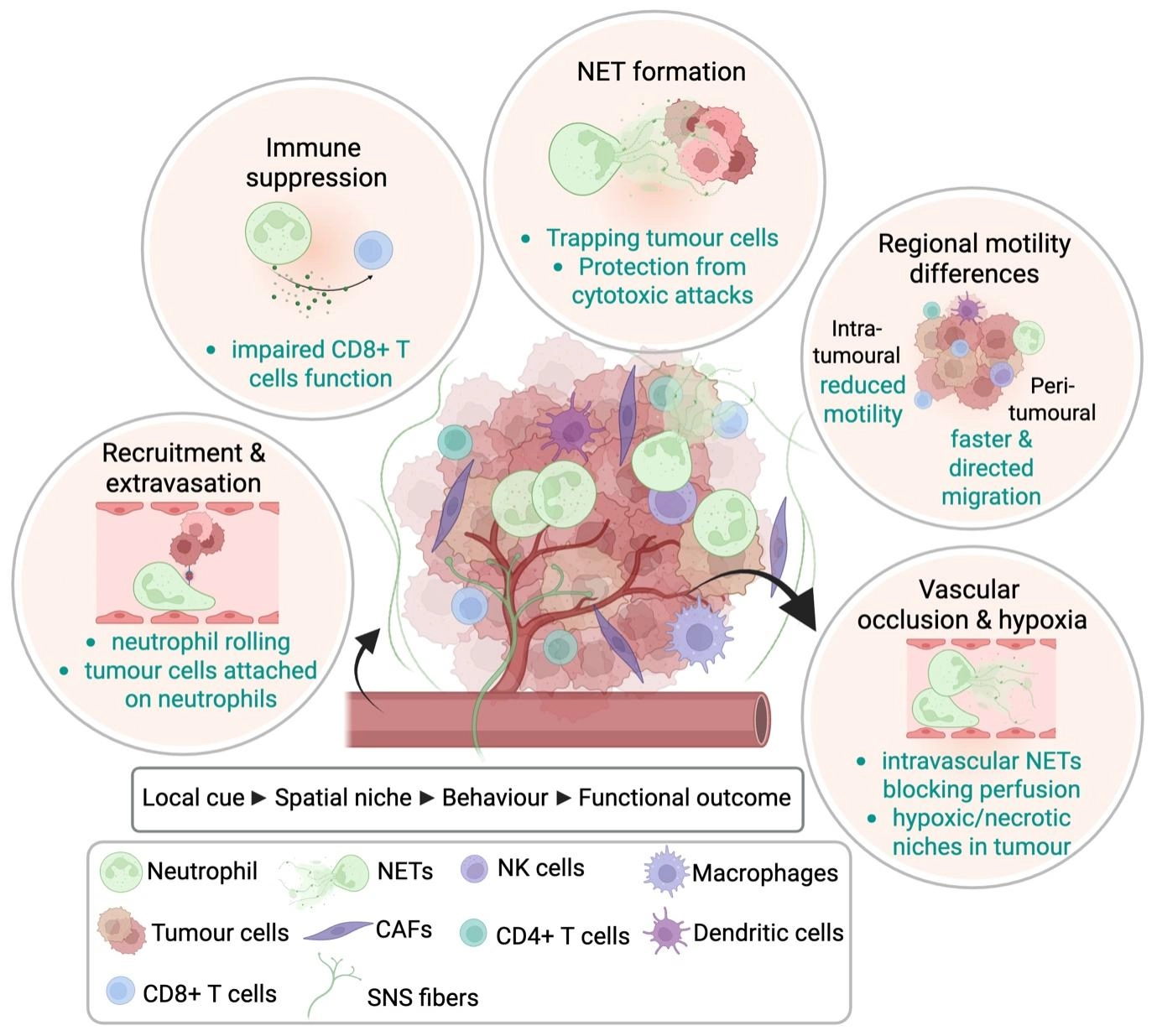
Figure 2. Recurring neutrophil behaviours in the TME revealed by intravital imaging. Real-time intravital imaging studies have shown that neutrophils display diverse and context-dependent behaviours across distinct tumour regions. In peri-tumoural blood vessels, neutrophils undergo recruitment, endothelial attachment, and extravasation into surrounding tissue. Within the peri-tumoural stroma and tumour core, neutrophils exhibit region-specific migratory behaviours, with differences in motility, directional persistence, and responsiveness to local cues. Intravital imaging has also revealed direct interactions between neutrophils and tumour cells, including support for tumour cell arrest and metastatic seeding in the vasculature. In some settings, neutrophils form NETs, which can protect tumour cells from CTLs and contribute to immune suppression. Recently, imaging studies have shown that intravascular neutrophil activities can impair tumour perfusion, generating hypoxic and necrotic regions within tumours. Together, these observations illustrate how neutrophil identity, spatial niche, and dynamic behaviour are linked to distinct functional outcomes in cancer. Created in BioRender. Devi, S. (2026) https://BioRender.com/0das480. TME: tumour microenvironment; NETs: neutrophil extracellular traps; NK: natural killer; CAFs: cancer-associated fibroblasts; SNS: sympathetic nervous system; CTLs: cytotoxic T lymphocytes.
Table 1. Overview of published intravital imaging studies examining neutrophils in tumour models. The table summarizes the tumour setting, anatomical site imaged, neutrophil-labelling approach, principal behaviour visualized and the key biological insight derived from live imaging.
| Tumour model | Image site | Detection strategy of neutrophils | Principal behaviour visualized | Key biological insight | AuthorYear [Ref] |
| Lewis lung carcinoma, H-59 cells | Liver sinusoids | Using white-light transillumination to visualise leukocytes; i.v. fluorescent conjugated anti-Gr-1 mAb | Neutrophil-tumour cell interaction in the microvasculature; early adhesion events | Neutrophils facilitate circulating tumour cell arrest and attachment within hepatic sinusoids, supporting early metastatic seeding | McDonald et al., 2009[59] Spicer et al., 2012[60] |
| Colon carcinoma, CT-26 cells | Primary tumour in skin flank | i.v. fluorescent conjugated anti-Ly6G Ab | Intra-tumoural motility, migration distance, velocity | Neutrophil dynamics within tumours can be quantified in vivo | Turk et al., 2018[65] |
| Hepatocellular cancer, Hep55.1C cells | Peri-tumoural vasculature and primary tumour in liver | c-FMS-eGFP reporter mice | Recruitment, extravasation, active migration into tumour tissue | Peri-tumoural vessels represent a major site of leukocyte recruitment before tissue infiltration and dissemination | Takeichi et al., 2010[66] |
| Murine oropharyngeal cancer, MOPC cells | Peri- and intra-tumoural regions in ear dermis | Catchup-IVMred reporter mice | Region-specific motility and CXCR2-dependent recruitment | Peri- and intra-tumoural neutrophils exhibit distinct migratory behaviours and differential dependence on CXCR2 | Sody et al., 2019[67] |
| Mouse Lewis lung carcinoma; melanoma, B16F10-3C cells; mammary carcinoma, AT-3; PDAC, KPC cells | Primary tumour in ear dermis | Catchup-IVMred and LysM-tdTomato reporter mice | Enhanced motility, tumour infiltration, tumour-cell deformation | Inflammatory stimulation can promote conversion toward a tumour-killing neutrophil phenotype associated with increased motility and infiltration | Yam et al., 2023[68] |
| Breast cancer, 4T-1 and 4T-07 cells | Lung metastatic niche | LysM-GFP reporter mice | Neutrophil accumulation and NET formation in metastatic sites | Neutrophils promote colonization of metastatic niches through NET-associated mechanisms | Park et al., 2016[69] |
| Breast cancer, 4T1 cells; HT29 tumour spheroid | Primary tumours in skin flank; spheroids | Neutrophils labelled with cell Tracker Deep Red. NET detection with Sytox Green | NET formation around tumour cells; interaction with CTLs | Tumour-derived CXCR1/2 agonists induce NETs that protect tumour cells from CD8+ T cell- and NK cell-mediated cytotoxicity | Teijera et al., 2020[70] |
| Mouse lewis lung carcinoma, LLC cells | Tumour vasculature and primary tumours in skin flank | LysM-GFP reporter mice | Intravascular NET formation, vascular occlusion, impaired perfusion | Neutrophils can block blood flow, promoting hypoxia, necrosis, and metastatic progression | Adrover et al., 2025[71] |
CXCR2: CXC chemokine receptor 2; NET: neutrophil extracellular trap; CTLs: cytotoxic T lymphocytes; NK: natural killer; CXCR1/2: CXC chemokine receptor 1 and 2; c-FMS: cellular FMS; eGFP: enhanced green fluorescent protein; PDAC: pancreatic ductal adenocarcinoma; LLC: lewis lung carcinoma.
Ng and colleagues showed that both mature and immature bone marrow–derived neutrophils infiltrate tumours and differentiate into distinct transcriptional states[28]. However, how neutrophils with these distinct characteristics behave dynamically within the TME remains largely unknown. Live imaging has shown that neutrophil motility is spatially regulated within the TME. In a murine model of oropharyngeal cancer, intra-tumoural neutrophils exhibited lower motility compared to peri-tumoural neutrophils, indicating that signals within the TME regulate their movement[67]. As tumours progressed, neutrophils further decreased both their velocity and directional persistence. Interestingly, recruitment of peri-tumoural, but not intra-tumoural, neutrophils was dependent on CXC chemokine receptor 2 (CXCR2), suggesting that distinct mechanisms govern neutrophil trafficking to different regions of the TME. Intravital imaging has revealed that peri-tumoural blood vessels are the major site of leukocyte recruitment, where, once recruited into the peri-tumoural tissues, extravasated leukocytes penetrate the tumour tissue, migrate actively, and disseminate within tumour tissues[66]. Notably, in a separate intravital imaging study, intra-tumoural injection of microbial bioparticles rapidly enhanced neutrophil motility, promoting their infiltration throughout the tumour[68]. Furthermore, regions of bioparticle-infused tumours with neutrophil infiltration exhibited increased tumour cell deformation, suggesting that the inflammatory microenvironment also enhanced neutrophil cytotoxic activity within the TME.
In metastatic settings, intravital imaging has provided direct evidence that neutrophils can facilitate tumour cell arrest, adhesion, and early colonization at distant sites, revealing real-time cell-cell interactions that are difficult to infer from endpoint analyses. In addition to the hepatic microcirculation studies described above[59,60,66], GFP+ neutrophils were observed to migrate to the lungs in a breast cancer metastasis model, where they promoted neutrophil extracellular traps (NETs) formation and facilitated colonization of new metastatic niches in the lungs[69]. Neutrophils can release DNA fiber networks comprised of histone and cytoplasmic granule proteins known as NETs in response to infection[29]. Importantly, intravital imaging revealed that TME-derived chemokines activate CXCR1 and CXCR2 on neutrophils, triggering NET formation that encases tumour cells and protects them from direct cytotoxic attacks by CD8+ T cells and NK cells[70].
Intravital imaging has further revealed that neutrophils can promote tumour progression through NET formation and intravascular activities that alter perfusion, generate hypoxia, and reshape the local TME. Recently, a tumour-associated neutrophil population was identified in multiple cancer mouse models that were able to form intravascular NETs that blocked blood flow inside the tumour vasculature, leading to regions with hypoxia and necrosis[71]. In a separate study, CCL3 produced by neutrophils supported the survival of neutrophils found in hypoxic tumour niches, thereby promoting tumour growth[72]. CTL migration slows in hypoxic tumour regions distant from blood vessels, highlighting the importance of oxygen availability for immune cell motility[46], yet the behaviour of neutrophils in hypoxic regions of the TME remains largely unexplored. Although still relatively understudied, intravital imaging also provides an opportunity to define how neutrophils influence the positioning and function of other immune cells in real time within tumours. Most current evidence suggests that tumour-associated neutrophils can shape anti-tumour immunity indirectly by modulating the local inflammatory milieu, restricting lymphocyte access, or interfering with cytotoxic immune function. However, these interactions remain incompletely resolved in vivo and represent an important area for future study.
Although neutrophils are now widely implicated in tumour progression and metastasis, only a handful of intravital imaging studies have captured their real-time behaviour within primary and metastatic tumours. Consequently, there remains a significant gap in our mechanistic understanding of how neutrophil recruitment and functional states are spatially and temporally regulated in vivo. A major challenge for the field is to determine how transcriptionally defined neutrophil states correspond to distinct behaviours and functions within the living TME. Future progress will likely depend on combining single-cell and spatially resolved profiling with live imaging approaches. For example, single cell RNA-sequencing datasets may help identify subset-enriched promoters, inducible transcriptional programs, or surface markers that can be exploited to develop more selective and reliable reporter systems for intravital imaging, such as the newly established CD101 reporter mice, to track mature neutrophils[64]. In parallel, correlative workflows that pair real-time imaging with post-imaging spatial transcriptomics, multiplex immunostaining, or tissue-clearing approaches may allow dynamically observed cells to be mapped back to their molecular states and local niches. Such strategies could help determine whether neutrophils occupying different tumour regions differ in activation state, lifespan, motility, suppressive capacity, or responsiveness to stromal and neural cues, thereby bridging the current gap between molecular heterogeneity and in vivo function.
6. Stromal and Neural Interactions in the Tumour Microenvironment
Emerging evidence indicates that the TME is shaped not only by cancer cells but also by stromal and neural components that influence immune cell behaviour. In particular, cancer-associated fibroblasts (CAFs) have been shown to modulate neutrophil recruitment, activation, and spatial distribution within tumours. CAFs secrete a wide repertoire of chemokines and cytokines, including CXCL12, CXCL1/2, IL-6, cardiotrophin-like cytokine factor 1 (CLCF1), and TGF-β, that not only attract circulating neutrophils but can also skew them toward a pro-tumourigenic N2 phenotype[73], enhance programmed death ligand 1 (PD-L1) expression[74], and promote NET formation[75], thereby suppressing local anti-tumour immunity and facilitating tumour progression. Beyond these chemotactic effects, CAFs modulate the architecture of the ECM and amplify inflammatory signalling cascades that further influence neutrophil trafficking and retention in tumour tissue. Parallel to stromal signalling, emerging evidence indicates that neural inputs also play an active role in shaping neutrophil recruitment, activation, and functional polarization within the TME.
Peripheral nerves, which innervate most tissues including tumours, are composed of distinct but complementary components, primarily sympathetic[76-78] and sensory fibres[79-81]. Sensory nerves, particularly nociceptive populations such as Nav1.8+ neurons, detect and transmit environmental cues including tissue damage, mechanical stress, and inflammation, enabling rapid communication between peripheral tissues and the central nervous system[82]. Using a Nav1.8 sensory neuron–specific designer receptors exclusively activated by designer drugs (DREADD) model, recent work has begun to uncover a functional link between sensory nerve activity and neutrophil dynamics within the TME[83]. In this approach, selective chemogenetic activation of Nav1.8+ nociceptive neurons led to a marked reduction in neutrophil infiltration into melanoma lesions, suggesting that sensory signalling can actively restrain neutrophil recruitment. This decrease in tumour-associated neutrophils was accompanied by a reduction in tumour growth, consistent with the established pro-tumourigenic roles of neutrophils in many cancer settings.
Sympathetic nerves, as part of the autonomic nervous system, regulate vascular tone and tissue homeostasis through the release of neurotransmitters such as norepinephrine, which bind to the adrenergic class of G-protein-coupled receptors that are widely expressed by many cell types[84]. Neutrophils predominantly express the β2-adrenergic receptor compared with other adrenergic receptor subtypes[85]. Studies in both in vivo and in vitro systems have established that adrenergic signalling influences neutrophil trafficking and shapes their effector functions[85-89]. For example, in a murine model mimicking chronic stress, β2-adrenergic receptor signalling drives persistent neutrophil recruitment to wounds via IL-6 upregulation, ultimately impairing tissue repair[89]. Notably, intravital imaging revealed that activation of local sympathetic neurons induced neutrophil mobilization from the bone marrow, promoting their egress and trafficking to peripheral sites[90]. However, whether sympathetic neural inputs directly influence neutrophil behaviour within tumours remains unexplored. Previous intravital imaging work demonstrates that sympathetic activation can cause lymphocytes to temporarily halt their migration in lymph nodes as a consequence of vascular constriction and hypoxia[23]. Tumours are frequently sympathetically innervated[76-78], raising the possibility that activation of the sympathetic nervous system (SNS) may influence neutrophil behaviour within tumours. Intravital imaging would serve as a powerful approach to investigate whether sympathetic stimulation alters neutrophil dynamics in the TME. Given that the TME is also hypoxic in nature, it remains unclear whether sympathetic signalling modulates neutrophil behaviour directly or indirectly through changes in local conditions, a question that has yet to be explored.
7. Translational Implication
The development of effective anti-tumour immunotherapies relies not only on identifying promising targets but also on understanding how these therapies function within the complex TME in vivo. While many mechanisms of action are inferred from in vitro studies or static analyses, these approaches often fail to capture the dynamic cellular interactions that underpin therapeutic efficacy. In this context, intravital imaging provides a powerful platform to directly visualize immune cell behaviour in real time within intact tissues. For example, intravital imaging studies in a melanoma mouse model revealed that combination immune checkpoint blockade enhances T cell motility more effectively than single-agent therapies[47]. Specifically, dual targeting of cytotoxic T lymphocyte-associated antigen-4 (CTLA-4) and PD-L1 increases T cell movement compared to PD-L1 blockade alone, while the addition of interleukin-2 (IL-2) further augments CTL migration in tumour models[91]. These findings suggest that combination strategies promote more dynamic T cell behaviour within tumours, which is associated with improved anti-tumour immune responses in vivo.
A range of cancer therapies, such as chemotherapy, radiotherapy, and immune checkpoint inhibitors (ICI), can influence both the abundance of circulating neutrophils and the recruitment or functional activity of tumour-associated neutrophils, thereby potentially affecting patient outcomes. Numerous preclinical and translational studies have reported that depletion of neutrophils, as well as pharmacological inhibition of CXCR2, can significantly enhance the efficacy of ICI therapies[92-98]. These approaches are thought to mitigate the immunosuppressive effects exerted by tumour-associated neutrophils, which often promote tumour progression by inhibiting CTL activity and fostering a pro-tumour microenvironment. By reducing neutrophil recruitment or function, CXCR2 blockade and Ly6G depletion help restore anti-tumour immune responses, thereby improving T cell infiltration and activation within tumours. Supporting this, an intravital imaging study demonstrated that CXCR2 inhibition effectively blocked neutrophil recruitment into tumours, however, this antagonistic effect was short-lived, with neutrophils eventually reappearing within the tumour core, albeit less prominently at the periphery[67]. Notably, neutrophils located within intra-tumoural versus peri-tumoural regions exhibited distinct behavioural patterns, suggesting spatial heterogeneity in their function and dynamics. While studies have shown that reducing neutrophil abundance or recruitment can enhance responses to immune checkpoint blockade, these strategies require careful interpretation. Ly6G-mediated depletion is primarily a mouse experimental tool used to interrogate neutrophil contribution in vivo and should not be viewed as a directly translatable therapeutic strategy. Similarly, although CXCR2 inhibition can reduce neutrophil recruitment and improve anti-tumour responses in some models, its effects may be incomplete or transient and are likely influenced by chemokine redundancy within the TME. Consistent with the immunosuppressive role of tumour-associated neutrophils, a recent study showed that the activation of CTLs during immunotherapy can drive interferon-γ (IFN-γ) production, which induces PD-L1 upregulation on neutrophils, thereby promoting a suppressive phenotype resembling aged and T3-like states that restrains cytotoxic activity[98]. Notably, anti-PD-L1 treatment in this setting was associated with neutrophil depletion from tumours and enhanced anti-tumour responses, further supporting neutrophil-mediated immune suppression as a therapeutically relevant axis. In addition, because neutrophils are essential for host defence and tissue homeostasis, broad suppression of their recruitment or survival may carry significant risks, including impaired antimicrobial immunity and altered inflammatory regulation. These limitations highlight the need for more selective approaches that reprogramme harmful neutrophil states while preserving protective functions.
Given that neutrophils are inherently short-lived and continuously replenished from the bone marrow, an important unresolved question is whether they can be durably reprogrammed at the level of granulopoiesis prior to their release into circulation. Emerging evidence from trained immunity and emergency myelopoiesis studies indicates that bone marrow progenitors can undergo durable functional reprogramming, raising the possibility that tumour-associated neutrophil responses may be shaped, at least in part, before tumour infiltration occurs[30,99]. In the context of cancer, this raises the possibility that systemic tumour-derived signals may shape neutrophil output at the level of haematopoietic progenitors, thereby biasing the phenotype and functional potential of cells before they enter the circulation or infiltrate tumours. Understanding whether trained or preconditioned neutrophil programs influence tumour defence may therefore open new opportunities to modulate neutrophil responses upstream of tumour entry.
Additionally, emerging evidence increasingly highlights the role of extrinsic systemic factors in shaping the TME, particularly the recruitment of neutrophils into tumours[73-75,83,97,100]. These findings suggest that the TME is not solely governed by local tumour-intrinsic mechanisms but is also dynamically regulated by broader physiological systems. Consequently, targeting such extrinsic pathways represents a promising therapeutic strategy, with the potential to reprogram the TME and enhance the efficacy of existing cancer treatments. In the context of combination with ICI therapy, it remains unclear how such interventions shape neutrophil behaviour within the TME. The observed differences between intra- and peri-tumoural neutrophils underscore the need to better understand how neutrophils are functionally reprogrammed in situ, and whether combination strategies can promote stable anti-tumour phenotypes. Future work should therefore focus on elucidating how targeting neutrophils alongside ICI influences their spatial distribution, functional states, and overall behaviour within the TME, with the goal of optimizing synergistic therapeutic outcomes. By enabling the observation of cell trafficking, interactions, and functional responses in situ, intravital imaging can refine and validate existing models of immunotherapy action, offering critical insights into how these treatments operate in vivo. Future intravital imaging studies should also consider biological variables such as sex and circadian timing, as both are increasingly recognised to influence neutrophil trafficking, activation, and immune function, with potential consequences for the interpretation of tumour-associated behaviours.
8. Conclusion
Emerging research has established that neutrophils exhibit remarkable plasticity and functional versatility within the TME. Given the inherent heterogeneity of the TME, neutrophils can adopt both pro- and anti-tumoural roles, often with conflicting effects on disease progression. In concert with broader host-derived influences, including systemic and neural cues, cancer can exploit neutrophils to support its own survival, immune evasion, and growth. These complexities underscore the urgent need for further research to delineate the conditions under which neutrophils can be therapeutically manipulated to enhance their anti-tumour functions while limiting their pro-tumoural activities.
Intravital imaging has provided unique insight into how neutrophils are recruited, positioned, and function within the living TME, revealing behaviours that are difficult to infer from static tissue analysis or molecular profiling alone. These studies have highlighted the spatial and temporal plasticity of neutrophils across tumour regions and metastatic sites, while also underscoring important gaps in our understanding of how transcriptionally defined neutrophil states relate to dynamic in vivo function. Although current imaging approaches remain limited by depth, field of view, motion artefacts, and the incomplete ability to distinguish neutrophil states, emerging combinations of intravital microscopy with functional biosensors, spatial transcriptomics, and high-dimensional tissue profiling offer a promising path forward. Future studies integrating these approaches will be important for defining when neutrophils support tumour progression, when they contribute to tumour control, and how these behaviours may be therapeutically reprogrammed. Consideration of biological variables such as sex and circadian timing will also be important, as both can influence neutrophil trafficking, activation, and immune function.
Acknowledgments
The author gratefully acknowledges Scott N Mueller for his valuable input in reviewing and discussions.
Authors contribution
The author contributed solely to the article.
Conflicts of interest
The author declares no conflicts of interest.
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Availability of data and materials
Not applicable.
Funding
This work was supported by funding from the National Health and Medical Research Council (NHMRC) Ideas Grant (GNT2038669) to SD.
Copyright
© 2026 The Author.
References
-
1. Pittet MJ, Weissleder R. Intravital imaging. Cell. 2011;147(5):983-991.[DOI]
-
3. Pittet MJ, Garris CS, Arlauckas SP, Weissleder R. Recording the wild lives of immune cells. Sci Immunol. 2018;3(27):eaaq0491.[DOI]
-
4. Condeelis J, Segall JE. Intravital imaging of cell movement in tumours. Nat Rev Cancer. 2003;3(12):921-930.[DOI]
-
7. Entenberg D, Oktay MH, Condeelis JS. Intravital imaging to study cancer progression and metastasis. Nat Rev Cancer. 2023;23(1):25-42.[DOI]
-
20. Peng X, Wang Y, Zhang J, Zhang Z, Qi S. Intravital imaging of the functions of immune cells in the tumor microenvironment during immunotherapy. Front Immunol. 2023;14:1288273.[DOI]
-
28. Ng MSF, Kwok I, Tan L, Shi C, Cerezo-Wallis D, Tan Y, et al. Deterministic reprogramming of neutrophils within tumors. Science. 2024;383(6679):eadf6493.[DOI]
-
29. Ballesteros I, Hidalgo A. The neutrophil collective. Cell. 2025;188(25):7019-7035.[DOI]
-
30. Cerezo-Wallis D, Rubio-Ponce A, Richter M, Pitino E, Kwok I, Marteletto G, et al. Architecture of the neutrophil compartment. Nature. 2026;649(8098):1003-1012.[DOI]
-
31. Wislez M, Rabbe N, Marchal J, Milleron B, Crestani B, Mayaud C, et al. Hepatocyte growth factor production by neutrophils infiltrating bronchioloalveolar subtype pulmonary adenocarcinoma: Role in tumor progression and death. Cancer Res. 2003;63(6):1405-1412.[PubMed]
-
33. Jensen HK, Donskov F, Marcussen N, Nordsmark M, Lundbeck F, von der Maase H. Presence of intratumoral neutrophils is an independent prognostic factor in localized renal cell carcinoma. J Clin Oncol. 2009;27(28):4709-4717.[DOI]
-
34. Trellakis S, Bruderek K, Dumitru CA, Gholaman H, Gu X, Bankfalvi A, et al. Polymorphonuclear granulocytes in human head and neck cancer: Enhanced inflammatory activity, modulation by cancer cells and expansion in advanced disease. Int J Cancer. 2011;129(9):2183-2193.[DOI]
-
37. Gabrilovich DI, Bronte V, Chen SH, Colombo MP, Ochoa A, Ostrand-Rosenberg S, et al. The terminology issue for myeloid-derived suppressor cells. Cancer Res. 2007;67(1):425.[DOI]
-
44. Gong Z, Li Q, Shi J, Li P, Hua L, Shultz LD, et al. Immunosuppressive reprogramming of neutrophils by lung mesenchymal cells promotes breast cancer metastasis. Sci Immunol. 2023;8(80):eadd5204.[DOI]
-
47. Lau D, Garçon F, Chandra A, Lechermann LM, Aloj L, Chilvers ER, et al. Intravital imaging of adoptive T-cell morphology, mobility and trafficking following immune checkpoint inhibition in a mouse melanoma model. Front Immunol. 2020;11:1514.[DOI]
-
51. Bougherara H, Mansuet-Lupo A, Alifano M, Ngô C, Damotte D, Le Frère-Belda MA, et al. Real-time imaging of resident T cells in human lung and ovarian carcinomas reveals how different tumor microenvironments control T lymphocyte migration. Front Immunol. 2015;6:500.[DOI]
-
52. Broz ML, Binnewies M, Boldajipour B, Nelson AE, Pollack JL, Erle DJ, et al. Dissecting the tumor myeloid compartment reveals rare activating antigen-presenting cells critical for T cell immunity. Cancer Cell. 2014;26(6):938.[DOI]
-
54. Ng M, Cerezo-Wallis D, Ng LG, Hidalgo A. Adaptations of neutrophils in cancer. Immunity. 2025;58(1):40-58.[DOI]
-
56. Devi S, Wang Y, Chew WK, Lima R, A-González N, Mattar CNZ, et al. Neutrophil mobilization via plerixafor-mediated CXCR4 inhibition arises from lung demargination and blockade of neutrophil homing to the bone marrow. J Exp Med. 2013;210(11):2321-2336.[DOI]
-
62. Faust N, Varas F, Kelly LM, Heck S, Graf T. Insertion of enhanced green fluorescent protein into the lysozyme gene creates mice with green fluorescent granulocytes and macrophages. Blood. 2000;96(2):719-726.[DOI]
-
64. Cao J, Yaw HP, Yi S, Zhou Y, Qin S, Wang Y, et al. Genetic tool for specific tracking of mature neutrophils. bioRxiv [Preprint]. 2026.[DOI]
-
79. Wei HK, Yu CD, Hu B, Zeng X, Ichise H, Li L, et al. Tumour-brain crosstalk restrains cancer immunity via a sensory-sympathetic axis. Nature. 2026;650:1007-1016.[DOI]
-
80. Barr J, Kruse A, Restaino A, Tulina N, Stuckelberger S, Vermeer S, et al. Intra-tumoral nerve-tracing in a novel syngeneic model of high-grade serous ovarian carcinoma. Cells. 2021;10(12):3491.[DOI]
-
82. Wood JN, Yan N, Huang J, Zhao J, Akopian A, Cox JJ, et al. Sensory neuron sodium channels as pain targets; from cocaine to Journavx (VX-548, suzetrigine). J Gen Physiol. 2025;157(4):e202513778.[DOI]
-
84. Mueller SN. Neural control of immune cell trafficking. J Exp Med. 2022;219(3):e20211604.[DOI]
-
85. Nicholls AJ, Wen SW, Hall P, Hickey MJ, Wong CHY. Activation of the sympathetic nervous system modulates neutrophil function. J Leukoc Biol. 2018;103(2):295-309.[DOI]
-
87. Trabold B, Gruber M, Fröhlich D. Functional and phenotypic changes in polymorphonuclear neutrophils induced by catecholamines. Scand Cardiovasc J. 2007;41(1):59-64.[DOI]
-
91. Ochoa MC, Sanchez-Gregorio S, de Andrea CE, Garasa S, Alvarez M, Olivera I, et al. Synergistic effects of combined immunotherapy strategies in a model of multifocal hepatocellular carcinoma. Cell Rep Med. 2023;4(4):101009.[DOI]
-
93. Kwak JW, Nguyen HQ, Camai A, Huffman GM, Mekvanich S, Kenney NN, et al. CXCR1/2 antagonism inhibits neutrophil function and not recruitment in cancer. OncoImmunology. 2024;13(1):2384674.[DOI]
-
97. Sun L, Clavijo PE, Robbins Y, Patel P, Friedman J, Greene S, et al. Inhibiting myeloid-derived suppressor cell trafficking enhances T cell immunotherapy. JCI Insight. 2019;4(7):e126853.[DOI]
-
98. Pei S, Pan Y, Liang H, Lei L, Lin Q, Mi J, et al. Neutrophil regulation of immunotherapy for cancer is controlled by type II interferon. Immunity. 2026.[DOI]
Copyright
© The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
Publisher’s Note
Science Exploration remains a neutral stance on jurisdictional claims in published
maps
and institutional affiliations. The views expressed in this article are solely those
of
the author(s) and do not reflect the opinions of the Editors or the publisher.
Share And Cite



Science Exploration Style
Devi S. Neutrophils in cancer: Insights from intravital imaging. Myeloid Cells. 2026;1:202607. https://doi.org/10.70401/mc.2026.0007
Tips
Copy completed.
Submit a Manuscript
Author Instructions
Cite this Article
Article Metrics
0
View
0
Download
Cited
Article Updates

- Abstract
- Keywords
- 1. Introduction
- 2. Intravital Imaging Modalities
- 3. Neutrophils: Good or Bad in Cancer?
- 4. Imaging the Tumour Microenvironment
- 5. Neutrophil Dynamics in Cancer Revealed by Imaging
- 6. Stromal and Neural Interactions in the Tumour Microenvironment
- 7. Translational Implication
- 8. Conclusion
- Acknowledgments
- Authors contribution
- Conflicts of interest
- Ethical approval
- Consent to participate
- Consent for publication
- Availability of data and materials
- Funding
- References
- Copyright
Science Exploration Style
Devi S. Neutrophils in cancer: Insights from intravital imaging. Myeloid Cells. 2026;1:202607. https://doi.org/10.70401/mc.2026.0007
copy
Share Link
copy