Advancing genetic enhancement of mesenchymal stem cells for combating aging and age-related diseases: Opportunities and challenges
Jinghui Lei
1

,
Si Wang
1,2,*
*Correspondence to:
Si Wang, Advanced Innovation Center for Human Brain Protection, National Clinical Research Center for Geriatric Disorders, Aging Translational Medicine Center, Beijing Municipal Geriatric Medical Research Center, Beijing Key Laboratory of Environment and Aging, Xuanwu Hospital Capital Medical University, Beijing 100053, China.
E-mail: wangsi@xwh.ccmu.edu.cn
Ageing Cancer Res Treat. 2026;3:202604. 10.70401/acrt.2026.0021
Received: January 13, 2026Accepted: May 22, 2026Published: May 22, 2026
Abstract
Mesenchymal stem cells (MSCs) hold substantial promise for treating aging and age-related diseases due to their regenerative and immunomodulatory properties. However, clinical applications remain limited by the poor survival and short retention of transplanted cells within the hostile aged microenvironment. Emerging genetic-engineering approaches, including viral vector-mediated genetic modification and genome-editing technologies such as clustered regularly interspaced short palindromic repeats/CRISPR-associated protein (CRISPR/Cas) systems, offer powerful strategies to enhance MSC resilience, functionality, and reparative capacity. Recent advances, such as the development of senescence-resistant mesenchymal progenitor cells (SRCs), demonstrate that precise modification of key longevity pathways can produce MSCs with superior stress resistance, reduced senescence, and improved regenerative performance in both rodent and non-human primate models. These findings highlight the potential of genetically enhanced MSCs as next-generation cellular therapeutics for precision interventions in aging. Nevertheless, challenges related to long-term safety, immunogenicity, off-target effects, and large-scale manufacturing must be addressed before clinical translation.
Keywords
Mesenchymal stem cells, genetic enhancement, cell therapy, aging, age-related diseases
1. Introduction
Aging manifests as a gradual deterioration of physiological function across multiple organ systems, and is a primary contributor to numerous age-associated pathologies such as cardiovascular diseases, diabetes mellitus, and neurodegenerative disease[1]. Amid global population aging, especially the rapid progression toward a deeply aged society in China, deciphering the biological basis of aging and establishing efficacious strategies for healthy aging have emerged as critical research imperatives in life medicine.
Adult stem cells (ASCs) are undifferentiated cells present in various tissues and organs of adult organisms, possessing self-renewal capacity and multipotent differentiation potential. They can differentiate into specific cell types of the tissues where they reside, thereby maintaining tissue homeostasis and repairing damage[2]. In the field of aging research, ASC senescence is considered an important mechanism of organismal aging. With age, the number of ASCs decreases and their functions decline, leading to reduced tissue repair capacity and increased susceptibility to diseases[3]. Whether supplementation with functionally intact stem cells can restore tissue repair capacity, remodel the aging microenvironment, and thereby delay aging and prevent age-related diseases remains an open scientific question. Of all stem cell types applicable to transplantation, mesenchymal stem cells (MSCs) are widely favored on account of their prominent biological features. These include broad tissue availability, multipotent differentiation potential, low immunogenicity, and intrinsic immunomodulatory activities[4,5].
In 2017, a landmark clinical trial conducted by the Miller School of Medicine at the University of Miami first evaluated the therapeutic effects of MSCs on frailty. The results showed that 30 elderly participants aged over 75 years who received MSC infusion exhibited improved physical function indicators such as 6-minute walk distance, and reduced levels of pro-inflammatory factors such as IL-6 in serum[6,7]. Furthermore, the research team developed Laromestrocel (Lomecel-BTM), an allogeneic human bone marrow-derived MSC preparation, and conducted a Phase 2b clinical trial involving 148 patients with frailty syndrome[8]. The results showed that compared with the placebo group, cell infusion achieved clinically meaningful, dose-dependent, and time-dependent improvements in the 6-minute walk test. These studies have laid a solid foundation for the clinical application of MSC-based interventions targeting aging. Nevertheless, in the context of aging and age-related diseases, organisms undergo detrimental microenvironmental alterations, including ischemia, hypoxia, increased oxidative stress, and chronic inflammation[9-11]. These aberrant changes impair the survival and retention of transplanted MSCs, thereby restricting their clinical translation and broad application. Merely increasing the number of transplanted cells to compensate for low survival rates will markedly raise therapeutic costs and may also induce adverse effects including cell aggregation and embolism. Therefore, developing key technologies for optimizing cell quality and improving the survival rate of transplanted cells, and enhancing the adaptability of transplanted cells in the harsh microenvironment of lesion sites are of great significance for promoting the application and clinical translation of MSC therapy.
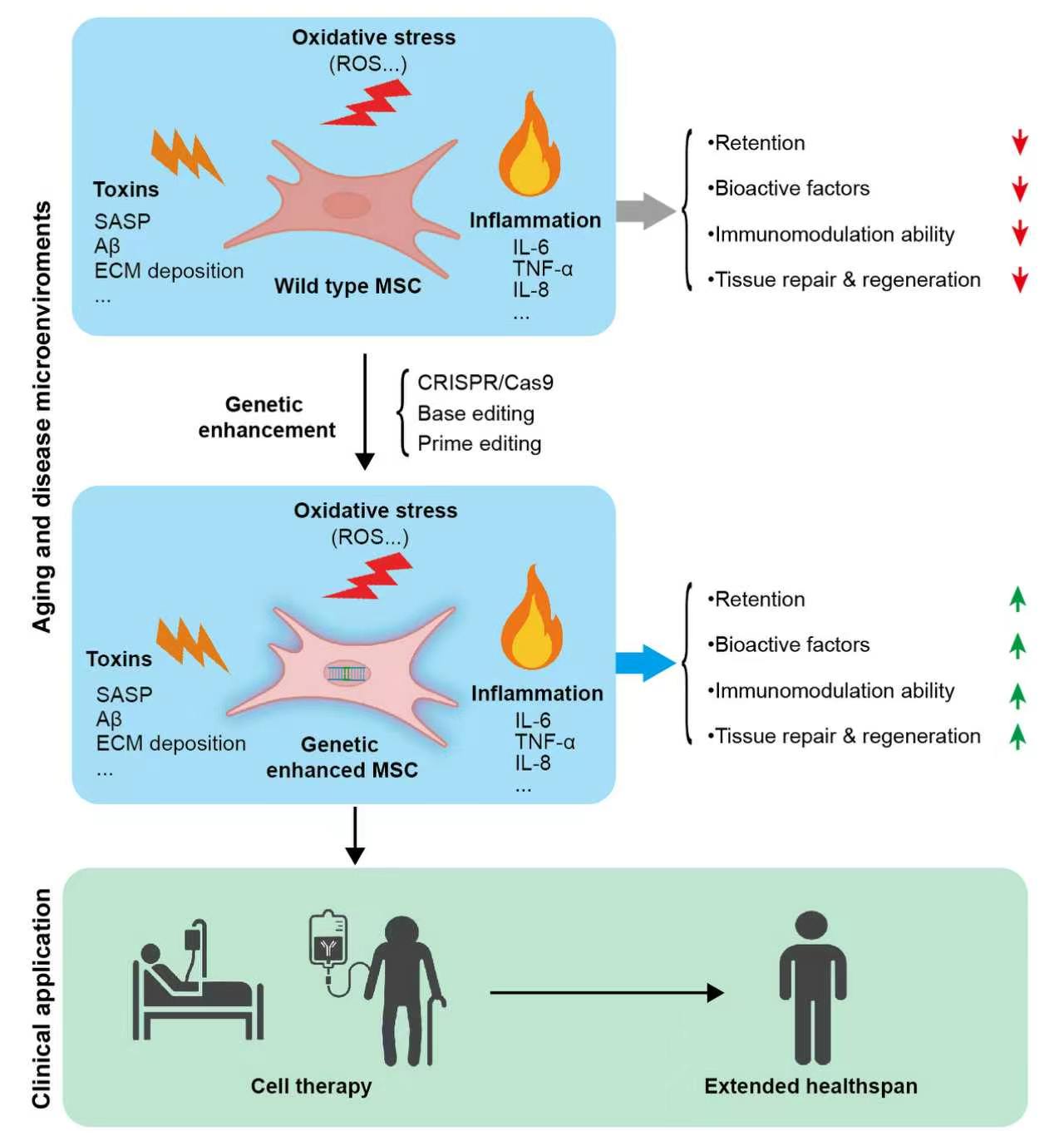
Previous studies have attempted to enhance cellular stress tolerance through various preconditioning strategies, such as exposing MSCs to in vitro stress conditions like hypoxia, hyperoxia, low-dose hydrogen peroxide, or serum deprivation, or co-transplanting MSCs with small-molecule drugs or biomaterials to improve their survival[12-14]. Collectively, these findings substantiate the theoretical feasibility of boosting the therapeutic performance of MSC transplantation. On the other hand, these preconditioning and co-transplantation strategies inevitably add complexity and pose potential risks. Major concerns encompass changes to cellular phenotypes and functions, non-specific activation of host tissues by pharmacological compounds, immune rejection elicited by biomaterials, and unknown persistence of preconditioning effects. These issues collectively result in variable outcomes and limit their translational potential for clinical application[15-17]. In recent years, in-depth insights into the biological properties of MSCs and aging-related mechanisms have offered new possibilities to overcome these bottlenecks, among which gene editing technology has emerged as a promising breakthrough approach. This article comprehensively discusses the opportunities and challenges of genetically engineered MSCs in aging intervention research, covering gene editing strategies, safety assessment, and advances in clinical translation (Figure 1).
{kind=link}
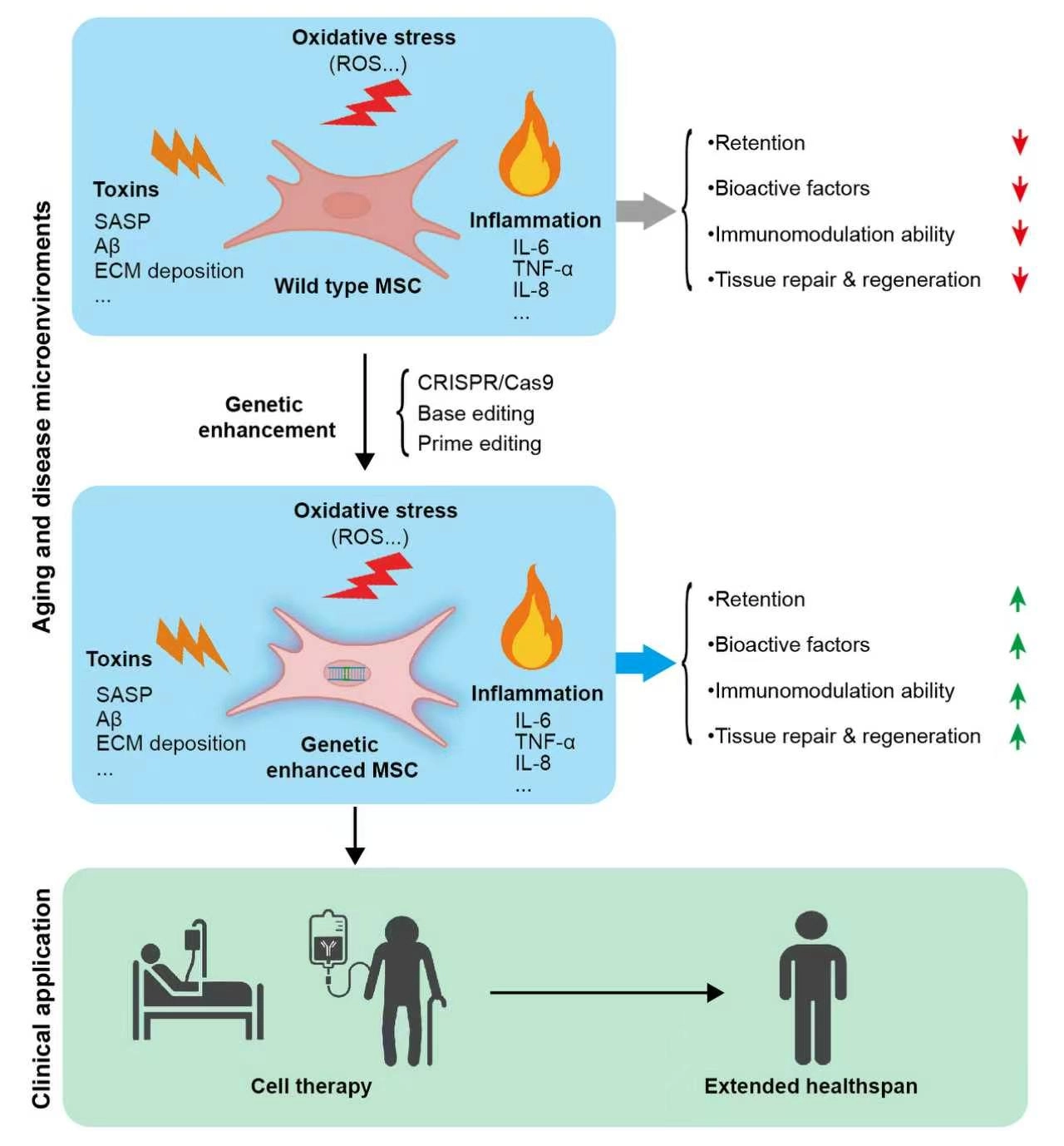
Figure 1. Schematic illustration of the functions and application prospects of genetically enhanced MSCs against aging- and disease-related microenvironmental stressors. Wild-type MSCs exhibit reduced retention, diminished secretion of bioactive factors, impaired immunomodulatory capacity, and weakened tissue repair and regeneration under harsh microenvironmental conditions such as oxidative stress, toxins, and chronic inflammation. Genetic enhancement endows MSCs with improved resilience to these stressors, resulting in increased cell retention, enhanced production of bioactive and trophic factors, strengthened immunomodulatory activity, and superior tissue repair and regenerative potential within aging and pathological microenvironments. MSCs: mesenchymal stem cells; ROS: Reactive oxygen species; SASP: Senescence-associated secretory phenotype; ECM: Extracellular matrix; TNF-α: Tumor necrosis factor-alpha; IL-6: interleukin-6; IL-8: interleukin-8; CRISPR/Cas9: clustered regularly interspaced short palindromic repeats/CRISPR-associated protein 9.
2. Non-site-specific Integration Gene Overexpression Strategies and Their Risks
Studies have shown that introducing specific genes into MSCs via viral vectors to achieve overexpression of beneficial molecules can enhance MSCs’ function, thereby realizing the transformation from “passive adaptation” toward the microenvironments to “active rewriting” of cell fate. As an example, overexpression of the anti-apoptotic gene Bcl-2 in rat MSCs diminished apoptosis by 32% under hypoxic stress and raised vascular endothelial growth factor release by more than 60%[18]. Transplantation of Bcl-2-overexpressing MSCs into ischemic myocardium reduced infarct size by 17% compared with the wild-type MSC group, accompanied by marked cardiac functional recovery[18]. C-X-C chemokine receptor type 4 (CXCR4) is the receptor for stromal cell-derived factor-1 (SDF-1) and plays an important role in MSC homing. Enhancing CXCR4 expression can enable MSCs to more effectively respond to the SDF-1 signal released by the injury site and promote their directional migration. Simultaneous overexpression of CXCR4 and IL-10 in MSCs markedly enhanced their homing capacity and anti-inflammatory potency in a mouse model of lipopolysaccharide (LPS)-induced local inflammation[19]. In addition, given that MSCs mainly release bioactive factors (such as growth factors and cytokines) through paracrine effects to promote repair, regulate immunity, and induce angiogenesis[20,21], researchers have overexpressed hepatocyte growth factor (HGF), insulin-like growth factor (IGF), and vascular endothelial growth factor (VEGF) in MSCs. Transplantation of edited MSCs into myocardial ischemic regions markedly enhanced vascular density, attenuated ischemic injury, and restored cardiac function[22-24]. In the nervous system, genetically modified MSCs overexpressing brain-derived neurotrophic factor (BDNF), glial cell line-derived neurotrophic factor (GDNF), exhibit potent neuroprotective properties compared with wild-type MSCs[25].
The above studies have verified the feasibility of enhancing MSC functions and therapeutic effects by targeting specific genes, providing new avenues for the development of cell therapy and regenerative medicine. However, traditional virus-mediated overexpression systems may lead to random insertion of genes, increasing the risk of tumorigenesis, which seriously restricts their clinical translational potential[25-27].
3. Site-specific Genetic Enhancement Strategies Based on Gene Editing
Gene editing technology offers the possibility to overcome the above bottlenecks. Unlike traditional random integration, gene editing can achieve precise modifications (insertion, deletion, replacement, etc.) at specific loci in the genome, thereby avoiding unintended insertion mutations. Combining gene editing technology with the research on aging regulatory mechanisms and longevity signaling pathways is expected to develop safer stem cell transplantation materials with geroprotective properties. The following two representative studies demonstrate the application of this strategy.
Nuclear factor erythroid 2-related factor 2 (NRF2) is a key transcription factor for resisting oxidative stress and achieving cell protection. A single-nucleotide substitution (c.245A>G; A245G) was introduced into the NRF2 locus of wild-type human embryonic stem cells (NRF2+/+ hESCs) via helper-dependent adenoviral vector (HDAdV)-mediated homologous recombination. This mutation altered the 82nd amino acid of NRF2 from glutamic acid to glycine (E82G), thereby enhancing NRF2 protein stability and transcriptional activity of its downstream target genes[28]. The data show that human mesenchymal stem cells differentiated from genetically modified human embryonic stem cells (NRF2AG/AG hESCs) display superior self-renewal and stress resistance in vitro. Upon orthotopic delivery into a mouse hindlimb ischemia model, these cells also achieve higher survival rates and robust pro-angiogenic function[28]. Notably, NRF2-enhanced MSCs are well tolerated in vivo and pose no tumorigenic risk[28].
Forkhead box O3 (FOXO3) is a well-recognized human longevity factor. Dual-site substitutions (T757G, T943G) were introduced into the FOXO3 locus via HDAdV-mediated homologous recombination. Such edits prevent phosphorylation-induced degradation, enabling sustained nuclear retention of FOXO3 and persistent activation of its geroprotective target genes[29]. Experimental results confirmed that FOXO3 genetic enhancement endowed MSCs with anti-senescence, resistance to harsh microenvironments and protection against oncogenic transformation, and these cells were therefore named “senescence-resistant human mesenchymal progenitor cells (SRCs)”[11,29]. In models of myocardial ischemia, hindlimb ischemia, cerebral ischemia and spinal cord injury, SRCs exhibit longer post-transplantation retention than wild-type MSCs, possess stronger tissue repair potency, and effectively facilitate vascular and neural tissue regeneration[29-32]. In addition, long-term in vivo experiments in aged macaques have proven that intravenous SRC infusion effectively ameliorates systemic aging manifestations, including cellular senescence, chronic inflammation and tissue degeneration, while improving cognitive performance, repairing tissue structure and reversing age-induced reproductive decline without causing adverse side effects[11].
Notably, SRC-derived exosomes (SRC-Exo) are capable of resetting the tissue aging clock in mice and alleviating senescence in multiple human cell lines. These vesicles exert stronger effects than exosomes isolated from wild-type MSCs (WTC-Exo). In another study, SRC-Exo partially reversed the senescent phenotype of epididymal epithelial cells[33]. In mouse models of spinal cord injury and cerebral ischemia, SRC-Exo also showed therapeutic potential by promoting neurogenesis and angiogenesis, with efficacy comparable to SRC transplantation[31,32]. Exosomal proteins, lipids and nucleic acids carry abundant information from “parent cells” and exert vital functions in the paracrine therapeutic effects of MSCs[34,35]. Consistent with this, proteomic and metabolomic analyses showed that SRC-Exo is rich in proteins related to anti-aging protection and metabolites that promote regeneration, providing a molecular basis for its functional advantages[11]. The above studies provide insights into the mechanism research of genetically enhanced stem cells in intervening aging and experimental support for the development of cell-free therapeutic strategies based on exosomes.
4. Future Expandable Targets and Gene Editing Tools
Genetic enhancement of MSCs via NRF2 and FOXO3 modifications not only generates novel cell candidates for aging intervention, but also provides new insights into overcoming the translational bottlenecks of stem cell therapy. Combining gene editing technology with mechanistic research on aging may facilitate the generation of superior stem cells capable of resisting the hostile aging microenvironment. For example, telomere shortening is closely related to cell senescence and serves as a biomolecular clock of cell age. Transient telomerase transfection or small molecule-mediated telomerase activation can effectively retard cellular senescence[36,37]. The Sirtuin family is also a key regulator of aging, playing a critical role in cell senescence, telomere maintenance, DNA repair, genome stability, and oxidative stress response[38]. Gene therapy based on SIRT2 or SIRT5 effectively delays aging in myocardium and skeletal muscle[39,40]. Both telomerase and Sirtuins family proteins are expected to show multi-level and multi-target application values in MSC function enhancement, and may provide a safe and efficient therapeutic paradigm for systemic aging reversal and ameliorating age-related diseases.
Equally important to the selection of intervention targets is enhancing the precision and safety of gene editing tools. At present, the most mature and widely used gene editing tool is the CRISPR/Cas system. The core principle of the CRISPR/Cas system is to use artificially designed guide RNA (gRNA) to accurately identify the target sequence of the genome, guide Cas nuclease to cleave DNA to form double-strand breaks, and cells complete gene knockout, insertion, or replacement through non-homologous end joining or homologous recombination repair[41]. Its advantages include precise targeting, high editing efficiency, simultaneous regulation of multiple genes, simple operation, and low cost, superior to traditional technologies such as Zinc Finger Nuclease (ZFN) and transcription activator-like effector nuclease (TALEN). The disadvantage is the risk of off-target effects and low delivery efficiency. Given the low gene editing efficiency of MSCs[42,43], the development of tailored gene editing strategies is essential to advance the translational application of genetically enhanced MSCs. In recent years, new gene editing technologies (such as base editing and prime editing) have developed rapidly. Characterized by the absence of double-strand DNA breaks, accurate single-base conversion, low off-target activity and superior safety, these editors hold great promise as core platforms for MSC genetic engineering. Base editing is a precise gene editing technology based on the CRISPR system, which can directly and irreversibly convert one base pair into another without cutting the DNA double strand[44]. Base editing has been applied to CD34+ hematopoietic stem and progenitor cells derived from beta-thalassemia patients to precisely edit specific bases at the HBG1/2 promoter locus. Infusion of the modified autologous hematopoietic stem cells (HSCs) resulted in sustained hematopoietic reconstitution in patients[45]. Although this study was mainly verified in HSCs, base editing is, in principle, also applicable to MSCs, providing feasibility for the genetic modification of MSCs and their use in disease treatment. Prime editing is another precise gene editing technology based on the CRISPR system. Compared with base editing technology, prime editing can achieve the replacement of any single base as well as the insertion and deletion of small fragments[46]. Base editing and prime editing represent the next generation of precise gene editing technologies, which achieve more flexible and controllable genome modification while avoiding risks related to double-strand breaks. Future advances combining novel delivery vectors, AI-assisted design, compact editing constructs, and recombinase systems will facilitate the integration of these technologies into stem cell research, and overcome bottlenecks hindering the clinical translation of functionally enhanced cell therapies.
5. Safety Evaluation: Existing Evidence and Potential Risks
From the perspective of clinical translation, in addition to efficacy evaluation, the safety of genetically enhanced MSCs needs to be considered more comprehensively. Existing studies on NRF2- and FOXO3-modified enhanced MSCs have confirmed the biosafety of these engineered cells. Tumorigenicity assessment in mouse models detected no evidence of malignant transformation of modified MSCs[28,29]. Moreover, the relevant FOXO3 studies further conducted multiple infusion in non-human primate aging models, with continuous monitoring of physiological indicators including body temperature, body weight, blood glucose, routine blood parameters and blood biochemistry, and no transplantation-associated adverse abnormalities were observed. Current safety evaluation systems lack unified detection benchmarks, with substantial discrepancies existing across different research protocols. In addition, mouse and primate studies are constrained by small sample sizes and limited observation durations, and large-sample, long-term follow-up data from aging host models remain scarce. Therefore, the current understanding of the safety of genetically enhanced MSCs still remains at an early stage.
More importantly, genetic modification based on gene editing (such as the CRISPR/Cas system) may introduce the following risks, most of which warrant further systematic investigation in future studies: (1) Unintended genetic alterations introduced by the editing process (off-target effects): CRISPR/Cas may cleave non-target sites, potentially inactivating tumor suppressor genes or activating proto-oncogenes. Even if next-generation tools reduce the off-target rate, low-frequency unintended edits may be amplified during long-term expansion of MSCs. Currently, most studies detect only a few predicted sites, lacking genome-wide off-target evaluation; (2) Unintended consequences at the targeted site (on-target but off-effect): Even when editing is successfully performed at the intended site, it may interfere with mRNA splicing, disrupt non-coding RNA function, or induce aberrant epigenetic modifications. This risk is distinct from off-target effects, arising from on-target but non-expected molecular events; (3) Immunogenicity and integration risks associated with delivery vectors: Due to the unsatisfactory editing efficiency of non-viral delivery in MSCs, viral vectors are widely used; however, they can trigger host immune responses or randomly integrate into the host genome, leading to insertion mutations (which also represent a form of genomic instability, but originate from the vector rather than the editing enzyme); (4) Tumorigenic potential inherent to overexpression of the intended gene itself: MSCs possess self-renewal capacity. Overexpressing anti-apoptotic genes or longevity genes, even in the absence of any off-target events or insertional mutations, theoretically carries a risk of uncontrolled proliferation or malignant transformation. Although no tumorigenesis has been observed in existing studies, this risk still requires long-term and multi-model validation.
In summary, future clinical translation needs to establish a more rigorous risk monitoring system, including genome-wide off-target detection, standardized physiological/pathological indicator monitoring, long term follow-up of large samples of aging hosts, and biosafety evaluation for integration mutations and tumorigenic transformation.
6. Current Status and Bottlenecks of Clinical Trials
To date, three clinical trials of genetically modified MSCs can be retrieved from ClinicalTrials.gov. The first one, hosted by Shanghai East Hospital, is to treat advanced colorectal cancer by overexpressing anti-tumor chemokines and co-stimulatory molecules in umbilical cord-derived MSCs (NCT06446050); the second one, hosted by Nuwacell Biotechnologies in Hangzhou, uses an undisclosed method to enhance the anti-fibrotic ability of iPSC-derived MSCs for interstitial lung disease (NCT06825169); the third one, from CHA University in South Korea, establishes bone marrow-derived MSCs overexpressing suicide genes for glioblastoma (NCT07143812). Notably, the indications of these trials are tumors or fibrotic diseases, not directly targeting aging or age-related diseases, and all at the Phase I clinical stage with no data published yet.
The above studies initially demonstrate the potential of genetic enhancement technology to accurately empower MSCs, but the global volume of relevant clinical research is still limited, and all are at the early exploration stage, with no available efficacy data and long-term safety evaluation. Critical safety hazards including off-target editing, unintended genomic alterations, immunogenicity, and long-term tumorigenic potential have yet to be thoroughly evaluated in clinical studies. Beyond general safety issues, in vivo application of these engineered cells faces persistent challenges: aberrant gene integration, suboptimal cell retention, and unstable differentiation, all of which impede further clinical translation. Further optimization of gene editing tools, establishment of standardized production workflows, and collection of long-term clinical data will fuel the development of regenerative and precision medicine, yielding innovative clinical approaches.
7. Conclusion and Perspective
In summary, gene editing enables the generation of potent, safer stem cells, presenting a viable approach to resolve bottlenecks in cell transplantation and advance the clinical translation of cell therapies. Nonetheless, several hurdles persist for the clinical implementation of genetically modified stem cell grafts. Despite proven in vivo safety of engineered MSCs in existing research, off-target editing and elicited host immune responses remain critical concerns. Standardized manufacturing and quality control pipelines are therefore essential to support their widespread use[47]. With ongoing mechanistic studies on aging regulation, longevity pathways and cellular stress responses, plus continuous improvements to gene editing and delivery platforms, genetically enhanced MSCs hold great promise as reliable therapeutics against aging and age-associated diseases, marking a major advance in regenerative medicine.
Acknowledgements
The authors declare that Doubao AI was only used for language polishing throughout manuscript compilation. No AI-based tools were adopted to develop the overall research framework or create original figures in this work.
Authors contribution
Wang S: Conceptualization, writing-review & editing, funding acquisition.
Lei J: Writing-original draft, writing-review & editing.
Conflicts of interest
Not applicable.
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Availability of data and materials
Not applicable.
Funding
This work was supported by the Program of the Beijing Natural Science Foundation (Grant No. Z240018), the National Natural Science Foundation of China (Grant Nos. 82471586 and 92468303)
Copyright
© The Author(s) 2026.
References
-
2. Mi L, Hu J, Li N, Gao J, Huo R, Peng X, et al. The mechanism of stem cell aging. Stem Cell Rev Rep. 2022;18(4):1281-1293.[DOI]
-
3. Goodell MA, Rando TA. Stem cells and healthy aging. Science. 2015;350(6265):1199-1204.[DOI]
-
5. Han X, Liao R, Li X, Zhang C, Huo S, Qin L, et al. Mesenchymal stem cells in treating human diseases: Molecular mechanisms and clinical studies. Sig Transduct Target Ther. 2025;10(1):262.[DOI]
-
18. Li W, Ma N, Ong LL, Nesselmann C, Klopsch C, Ladilov Y, et al. Bcl-2 engineered MSCs inhibited apoptosis and improved heart function. Stem Cells. 2007;25(8):2118-2127.[DOI]
-
19. Hervás-Salcedo R, Fernández-García M, Hernando-Rodríguez M, Quintana-Bustamante O, Segovia JC, Alvarez-Silva M, et al. Enhanced anti-inflammatory effects of mesenchymal stromal cells mediated by the transient ectopic expression of CXCR4 and IL10. Stem Cell Res Ther. 2021;12(1):124.
-
22. Deuse T, Peter C, Fedak PW, Doyle T, Reichenspurner H, Zimmermann WH, et al. Hepatocyte growth factor or vascular endothelial growth factor gene transfer maximizes mesenchymal stem cell-based myocardial salvage after acute myocardial infarction. Circulation. 2009;120(11 Suppl):S247-S254.
-
27. Nasiri H, Manoochehrabadi T, Eskandari F, Majidi J, Gholipourmalekabadi M. Genetic modification of mesenchymal stem cells (MSCs): Novel strategy to expand their naïve applications in critical illness. Mol Biol Rep. 2025;52(1):501.[DOI]
-
38. Lee SH, Lee JH, Lee HY, Min KJ. Sirtuin signaling in cellular senescence and aging. BMB Rep. 2019;52(1):24-34.[DOI]
-
39. Ye Y, Yang K, Liu H, Yu Y, Song M, Huang D, et al. SIRT2 counteracts primate cardiac aging via deacetylation of STAT3 that silences CDKN2B. Nat Aging. 2023;3(10):1269-1287.[DOI]
-
40. Zhao Q, Jing Y, Jiang X, Zhang X, Liu F, Huang H, et al. SIRT5 safeguards against primate skeletal muscle ageing via desuccinylation of TBK1. Nat Metab. 2025;7(3):556-573.[DOI]
-
43. Dashti M, Mohammaddust Sarab M, Shad F, Dehnavi S. CRISPR-mediated engineering of mesenchymal stromal/stem cells: A summary of recent progress in immunological applications for regenerative medicine and cancer therapy. Stem Cell Res Ther. 2025;16(1):592.[DOI]
-
44. Komor AC, Kim YB, Packer MS, Zuris JA, Liu DR. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature. 2016;533(7603):420-424.[DOI]
-
45. Lai Y, Liu R, Wang L, Ma XK, Li Y, Yang G, et al. Clinical application of base editing for treating β-thalassaemia. Nature. 2026;653(8115):923-932.[DOI]
Copyright
© The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
Publisher’s Note
Science Exploration remains a neutral stance on jurisdictional claims in published
maps
and institutional affiliations. The views expressed in this article are solely those
of
the author(s) and do not reflect the opinions of the Editors or the publisher.
Share And Cite



Science Exploration Style
Lei J, Wang S. Advancing genetic enhancement of mesenchymal stem cells for combating aging and age-related diseases: Opportunities and challenges. Ageing Cancer Res Treat. 2026;3:202604. https://doi.org/10.70401/acrt.2026.0021
Tips
Copy completed.
Submit a Manuscript
Author Instructions
Cite this Article
Article Metrics
0
View
0
Download
Cited
Article Updates

- Abstract
- Keywords
- 1. Introduction
- 2. Non-site-specific Integration Gene Overexpression Strategies and Their Risks
- 3. Site-specific Genetic Enhancement Strategies Based on Gene Editing
- 4. Future Expandable Targets and Gene Editing Tools
- 5. Safety Evaluation: Existing Evidence and Potential Risks
- 6. Current Status and Bottlenecks of Clinical Trials
- 7. Conclusion and Perspective
- Acknowledgements
- Authors contribution
- Conflicts of interest
- Ethical approval
- Consent to participate
- Consent for publication
- Availability of data and materials
- Funding
- References
- Copyright
Science Exploration Style
Lei J, Wang S. Advancing genetic enhancement of mesenchymal stem cells for combating aging and age-related diseases: Opportunities and challenges. Ageing Cancer Res Treat. 2026;3:202604. https://doi.org/10.70401/acrt.2026.0021
copy
Share Link
copy