Nanomaterial-based optimization of chemo-radiotherapy for precisely overcoming PD-L1 resistance in the tumor immune microenvironment
Bassam Lutf
1,2
,
Yuan Li
1,*
,
Jianliang Shen
2,3,*

*Correspondence to:
Yuan Li, Department of Urology, The Second Xiangya Hospital, Central South University, Changsha 410011, Hunan, China.
E-mail: yuanlixy@csu.edu.cn
Jianliang Shen, National Engineering Research Center of Ophthalmology and Optometry, Eye Hospital, Wenzhou 325027, Zhejiang, China. E-mail: shenjl@wiucas.as.cn
Jianliang Shen, National Engineering Research Center of Ophthalmology and Optometry, Eye Hospital, Wenzhou 325027, Zhejiang, China. E-mail: shenjl@wiucas.as.cn
BME Horiz. 2026;4:202617. 10.70401/bmeh.2026.0031
Received: March 27, 2026Accepted: July 06, 2026Published: July 07, 2026
Abstract
Immune checkpoint inhibitors, like PD-1/PD-L1 blockers, have revolutionized cancer treatment in recent years. However, these therapies still face many problems, such as low objective response rates and drug resistance, making clinical treatment very difficult. Numerous studies have demonstrated that the heterogeneity of the tumor microenvironment (TME), including various subtypes such as “cold tumors”, “hot tumors”, and “immune-rejecting tumors”, as well as complex immunosuppressive mechanisms, is a major factor leading to resistance to immunotherapy. Chemotherapy and radiotherapy can aid in treatment by inducing immunogenic cell death, but they often lead to PD-L1 upregulation, which causes treatment effects to vary over time and space and reduces their efficacy. Given their excellent biocompatibility, drug delivery capacity, and responsiveness to environmental factors, nanomaterials offer a novel opportunity to address this challenge. This review explores sequential nanoregulation strategies based on different types of TME, describes how functionalized nanocarriers, such as stimulus-responsive polymers, liposomes, and biomimetic nanoparticles, can be used to detect changes in the microenvironment, like pH, enzymes, and reactive oxygen species, and to precisely deliver drugs when needed. This will allow researchers to target key points of immune activation, thereby blocking immune checkpoint signaling. These specific strategies, which include antigen release, immune system activation, and blocking interventions, not only offer new approaches for overcoming PD-L1 resistance but also shed light on the challenges and opportunities associated with applying these strategies in clinical practice.
Keywords
Tumor microenvironment, PD-L1/PD-1, chemotherapy, radiotherapy, nanoparticles
1. Introduction
Immune checkpoint inhibitors (ICIs), especially PD-1/PD-L1 inhibitors, have revolutionized cancer treatment in recent years[1]. These therapies have demonstrated remarkable clinical efficacy in various advanced malignant tumors by mitigating the immunosuppressive effects of tumors on T cells. Although immunotherapy has made significant progress, it still faces many unresolved challenges. For example, only 15% to 30% of patients with solid tumors respond to immunotherapy[2,3]. Many patients either exhibit primary resistance from the outset or rapidly develop acquired resistance after a brief period of efficacy. The low response rate has hindered the widespread adoption of immunotherapy, so researchers are investigating why tumors develop resistance to it. This limited response rate varies significantly across cancer types, with higher responsiveness observed in melanoma and non-small cell lung cancer (NSCLC), while tumors such as pancreatic cancer, prostate cancer, and colorectal cancer generally exhibit strong primary resistance to PD-1/PD-L1 blockade[4-9]. These differences highlight the heterogeneity of PD-L1 resistance across clinical settings and emphasize the need for tumor-specific therapeutic strategies.
Accumulating evidence suggests that resistance to ICIs cannot be fully explained by tumor-intrinsic genetic alterations alone. Increasing attention has therefore been directed toward the tumor microenvironment (TME), which plays a central role in shaping antitumor immune responses[10]. The TME is usually divided into three types based on how well immune cells can get into the tumor: hot tumors, cold tumors, and immune-rejecting tumors. “Hot” tumors are more likely to respond to ICIs than “cold” tumors and “immune-rejecting” tumors[11].
Furthermore, regarding the TME, traditional treatments like chemotherapy and radiotherapy (RT) have their own problems when used with immunotherapy. A moderate dose of chemotherapy drugs can kill tumor cells and release “danger signals” (damage-associated molecular patterns, DAMPs), thereby “activating” the immune system and helping it recognize tumor antigens. On the other hand, radiation therapy utilizes the cGAS-STING pathway to convert DNA damage into an immune signal, thereby triggering an immune response[12]. However, studies have shown that chemotherapy drugs may enhance the expression of PD-L1 on the surface of tumor cells by activating inflammation-related signaling pathways, including NF-κB. This may help tumor cells protect themselves from attack by the immune system[13]. Radiation therapy also increases PD-L1 expression by releasing inflammatory mediators such as IFN-γ. This triggers feedback defense mechanisms involving immune checkpoints, leading to reduced efficacy of immunotherapy with long-term use[14,15].
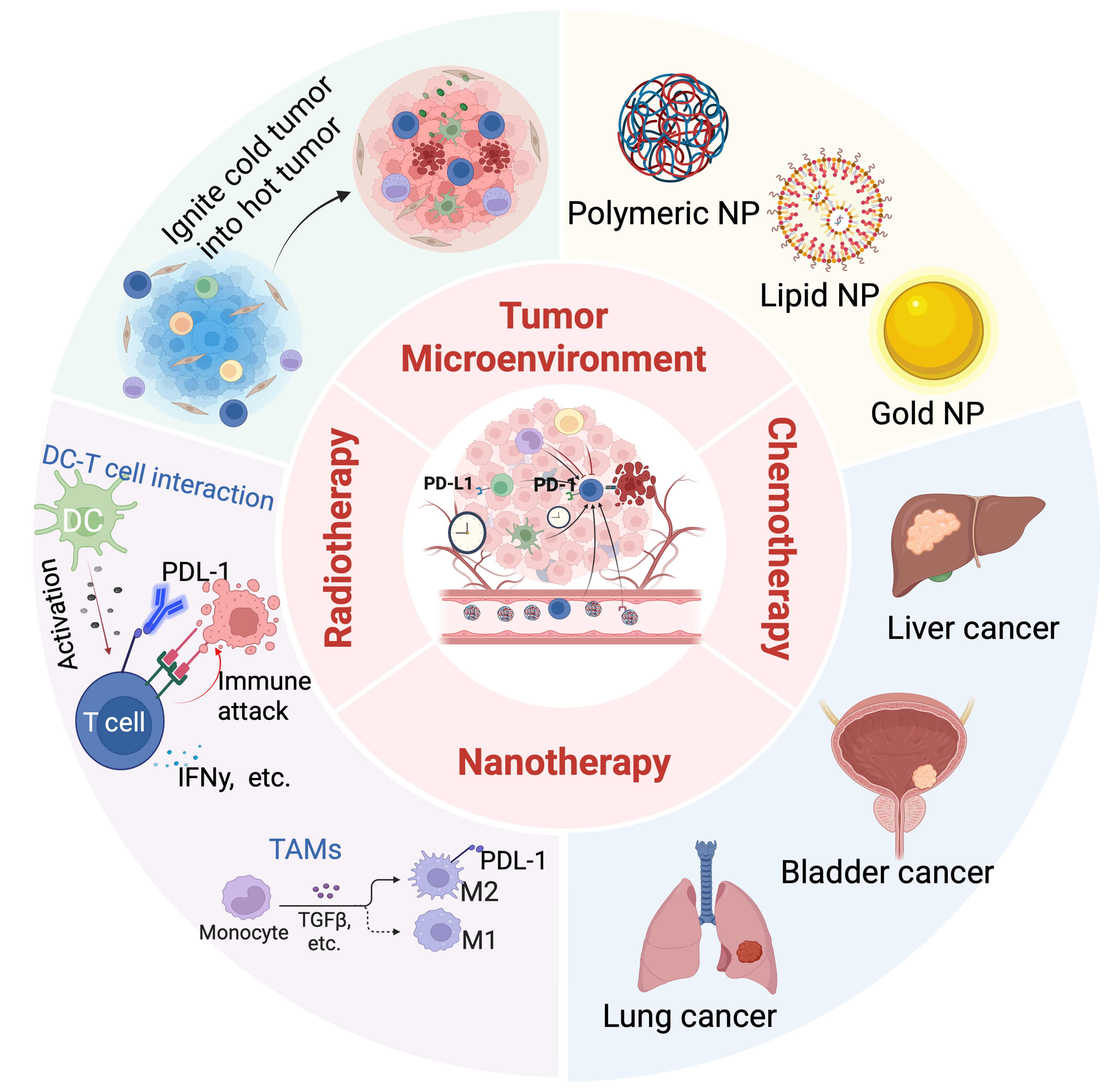
Although both chemotherapy and RT can enhance the function of the immune system, when used in combination with immunotherapy, they often encounter difficulties in clinical application due to the mismatch between the timing and site of treatment. Nanotechnology has proven in recent years to be able to solve the problem of synergistic therapy regimens that were previously impossible. By developing nanoplatforms that respond to their environment, researchers are able to precisely control when and where drugs are released. This significantly enhances the synergistic effects of different types of treatments. Additionally, controllable nanocarriers can release ICIs at the appropriate time, such as when chemotherapy and radiation therapy cause PD-L1 upregulation to compensate for the loss of immune cells. This precise control over time and space creates a perfect synergy between antigen release, immune system activation, and therapeutic intervention. This review summarizes nanomedicine strategies based on TME types and explores the potential benefits and challenges of combining these strategies with chemotherapy, radiation therapy, and immunotherapy in overcoming PD-L1 resistance and enhancing systemic immune responses (Figure 1).
{kind=link}
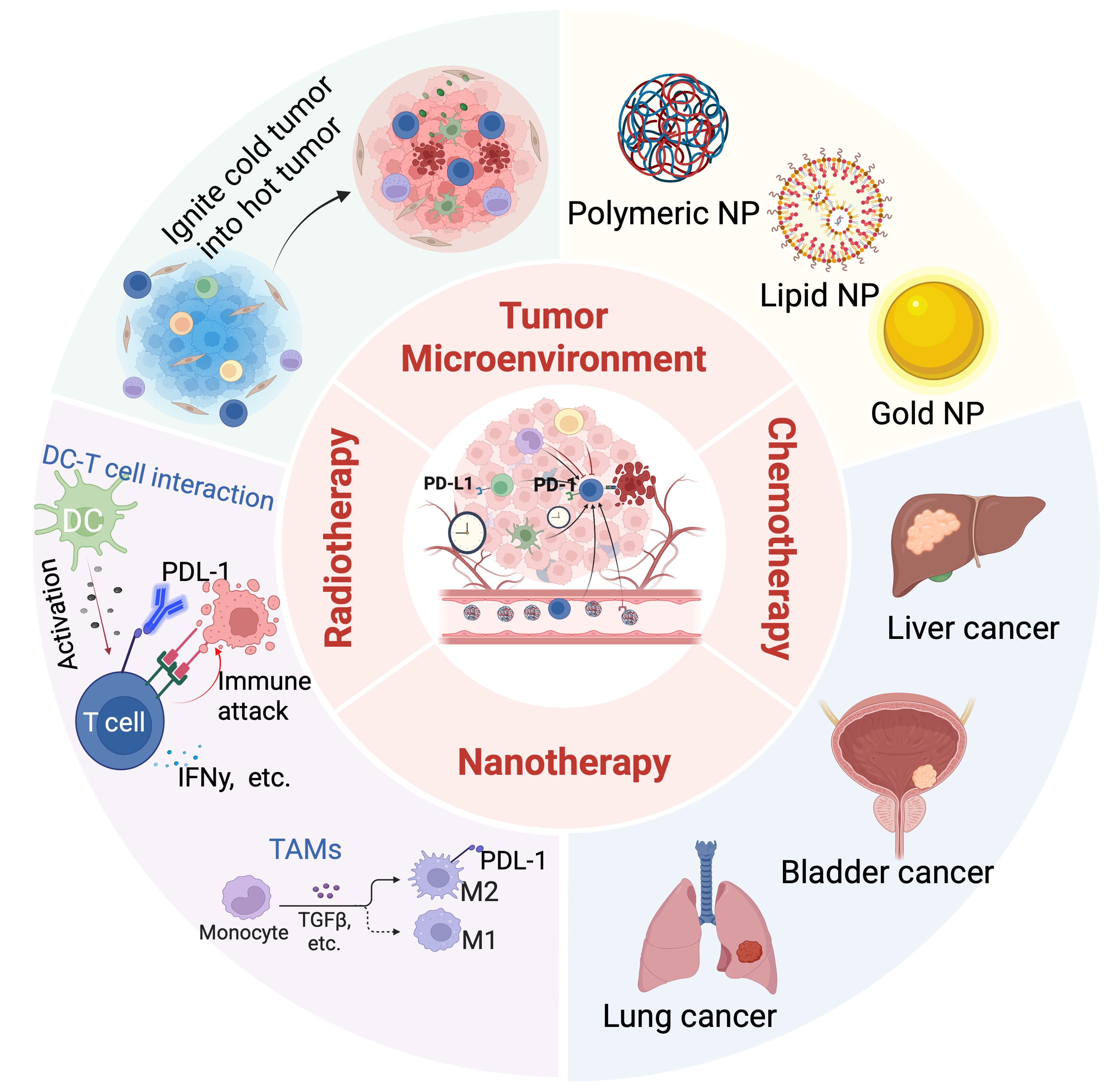
Figure 1. A schematic diagram illustrating common nanomaterials and their applications in cancer treatment, including RT, chemotherapy, and nanotherapy. Created in BioRender. Lutf, B. (2026) https://BioRender.com/ixmdm2i. RT: radiotherapy; NP: nanoparticle; IFN: interferon; TGF: transforming growth factor; TAMs: tumor-associated macrophages; DC: dendritic cell.
2. Molecular Mechanisms of PD-L1 Resistance and Heterogeneity of TME
PD-1/PD-L1 immune checkpoint blockade (ICB) has emerged as a standard therapeutic strategy for multiple malignancies. However, substantial molecular differences between “hot” and “cold” tumors critically determine immunotherapy responsiveness[11]. Elucidating the regulatory networks underlying PD-L1 expression across distinct TME contexts is therefore essential for understanding immune escape and guiding therapeutic design.
2.1 Molecular mechanisms regulating PD-L1 expression
PD-L1 expression is regulated through both intrinsic genetic programs and adaptive signaling responses. Intrinsic PD-L1 expression is mainly driven by oncogenic and epigenetic alterations, independent of exogenous stimuli, and is closely associated with the baseline immune phenotype of the TME. For example, PTEN loss or MYC amplification can sustain PD-L1 expression via constitutive oncogenic signaling[16-19].
In contrast, adaptive PD-L1 expression is induced by external stressors such as immune pressure and therapeutic interventions, including chemoradiotherapy (CRT). This reflects a dynamic “adaptive immune resistance” mechanism that enhances tumor immune evasion under treatment stress[14,20-23].
2.1.1 NF-κB in transcriptional regulation of PD-L1
NF-κB is a central transcription factor in inflammatory and stress responses and is considered a key positive regulatory hub for PD-L1 expression in tumors[13,24]. It can directly bind to the PD-L1 promoter to promote transcription and can also indirectly enhance its expression through multiple signaling networks, thereby driving immune escape within the TME[13].
In tumor cells, NF-κB integrates oncogenic and inflammatory signaling pathways, positioning it as a central node linking tumor progression and immune evasion. Chemotherapy-induced stress can activate NF-κB p65, thereby promoting PD-L1 transcriptional upregulation. This process is closely associated with the DNA damage response (DDR) signaling pathway and checkpoint regulation and may vary depending on the genetic background of tumors[13,25,26].
Furthermore, the DDR serves as a key upstream mechanism linking CRT to the NF-κB/PD-L1 axis. The ataxia telangiectasia mutated/ataxia telangiectasia and RAD3-related signaling pathway can activate NF-κB and the STAT1/STAT3/IRF1 pathways, synergistically enhancing PD-L1 expression[15,27]. Additionally, NF-κB can participate in DNA repair processes and enhance tumor cell survival, further promoting treatment resistance[28].
2.1.2 The cGAS-STING-IFN pathway
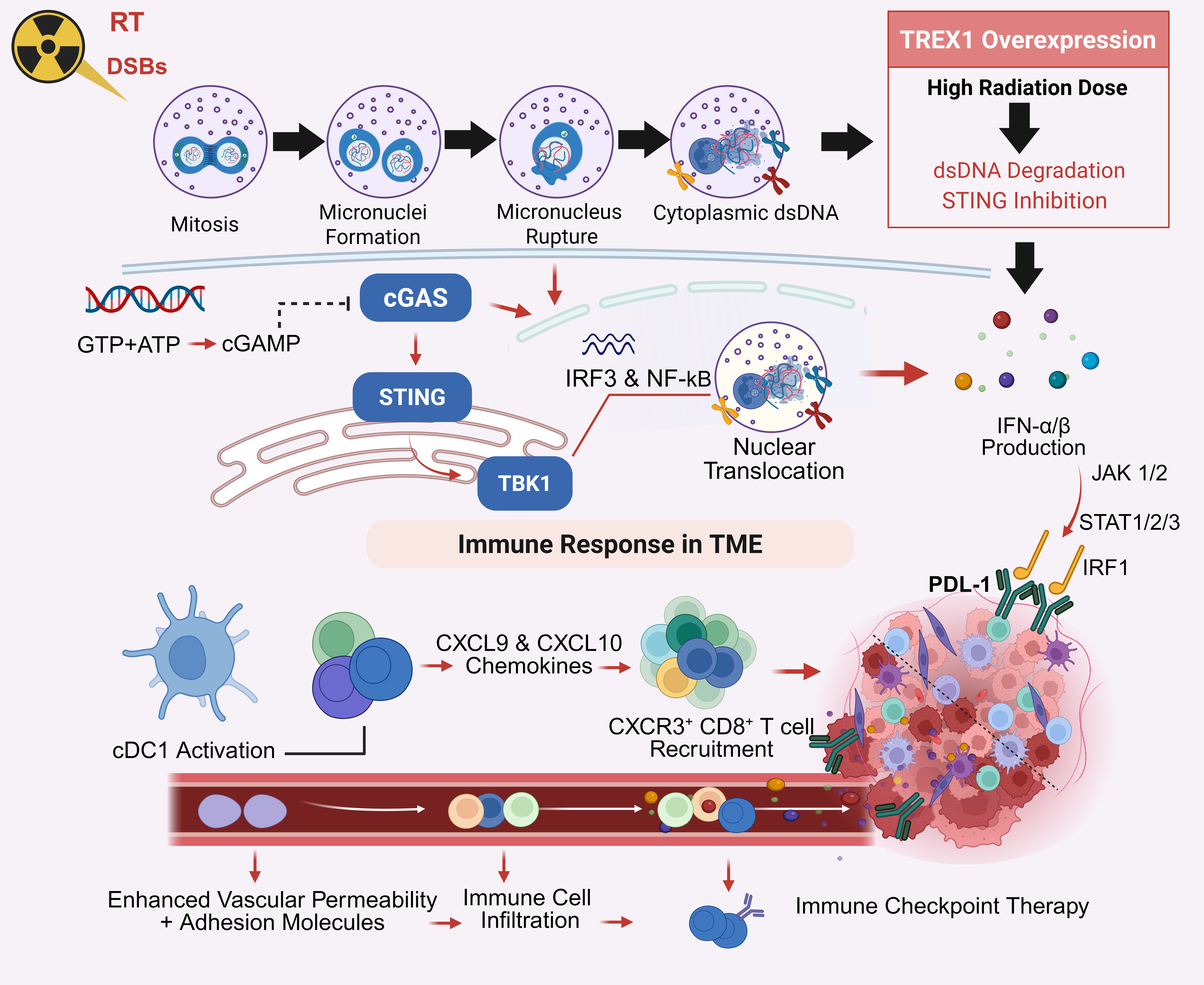
The cGAS-STING pathway is a critical innate immune signaling axis that mediates DNA damage sensing and plays a key role in RT-induced antitumor immune activation (Figure 2)[29,30]. DNA double-strand breaks (DSBs) generated by RT lead to cytoplasmic dsDNA accumulation, which is recognized by cGAS and triggers the STING-TBK1 cascade, subsequently inducing IRF3 and NF-κB activation and promoting the secretion of type I interferons and various pro-inflammatory cytokines[20].
{kind=link}
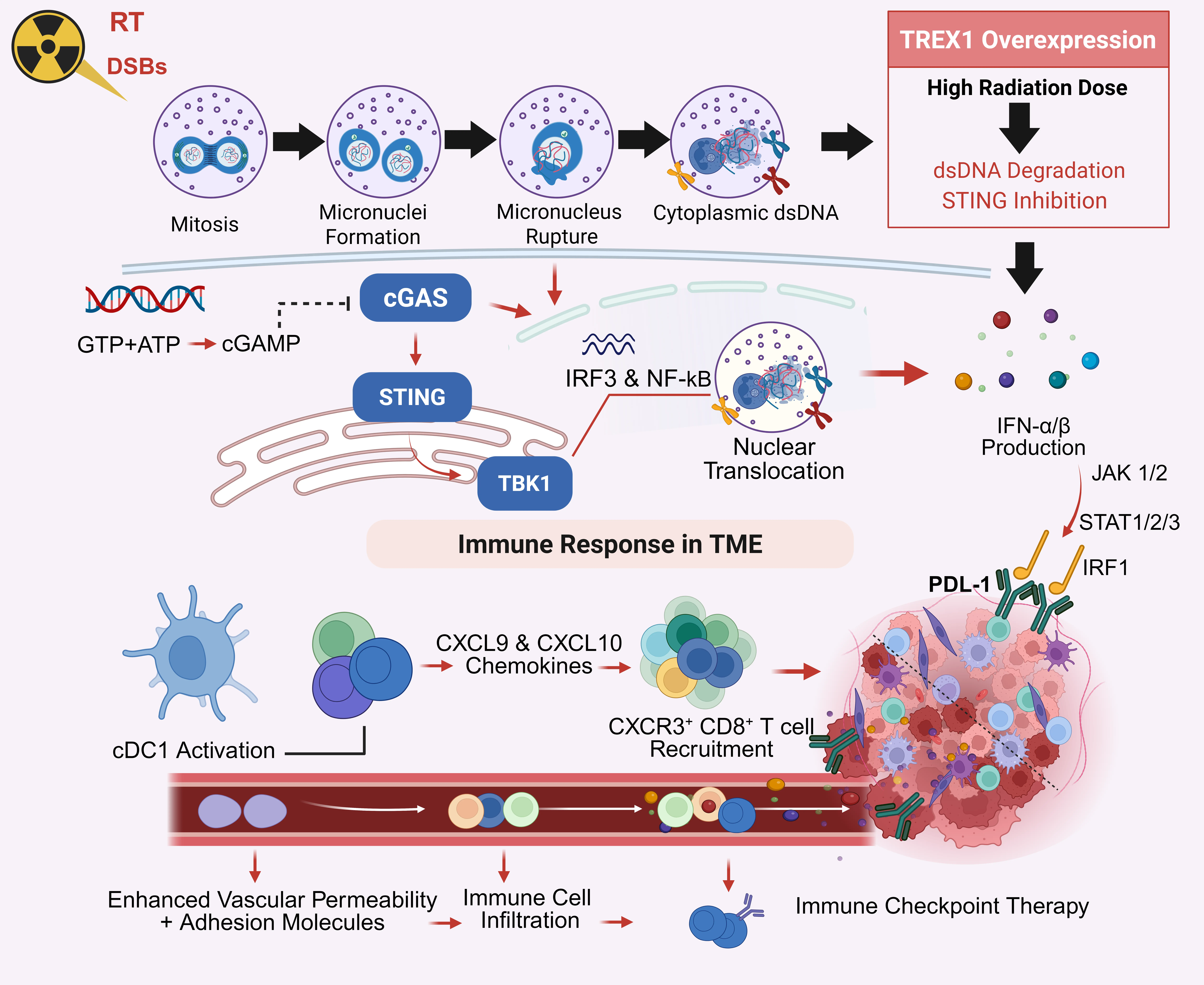
Figure 2. Schematic illustration of RT-induced cGAS-STING signaling and immune activation in the tumor microenvironment. RT induces DNA DSBs, leading to micronuclei formation and subsequent micronuclear rupture, which releases dsDNA into the cytoplasm. Cytosolic dsDNA activates the cGAS-STING-TBK1 signaling pathway, promoting IRF3 and NF-κB activation and inducing type I interferon production. Type I interferon signaling further activates the JAK1/2-STAT1/2/3-IRF1 axis, contributing to PD-L1 upregulation in the tumor microenvironment. These events enhance DC activation, CXCL9/CXCL10-mediated CXCR3+ CD8+ T-cell recruitment, vascular permeability, immune-cell infiltration, and the response to immune checkpoint therapy. In contrast, high-dose radiation may induce TREX1 overexpression, resulting in cytosolic dsDNA degradation and inhibition of STING signaling. Created in BioRender. Lutf, B. (2026) https://BioRender.com/qx3l09z. RT: radiotherapy; cGAS: cyclic GMP-AMP synthase; STING: stimulator of interferon genes; DSBs: double-strand breaks; dsDNA: double-stranded DNA; TBK: TANK-binding kinase; IRF: interferon regulatory factor; NF-κB: nuclear factor kappa B; STAT: signal transducer and activator of transcription; DC: dendritic cell; CXCL: C-X-C motif chemokine ligand; CXCR: C-X-C motif chemokine receptor; TREX: three-prime repair exonuclease.
This pathway can exert context-dependent effects within the TME. It may enhance antitumor immunity by promoting antigen presentation and dendritic cell (DC) activation, while also contributing to adaptive immune resistance under sustained activation conditions[30-34]. The downstream IFN-JAK-STAT axis further links STING activation to PD-L1 transcription. STAT1/STAT2/STAT3 activation via JAK1/2 induces IRF1-mediated PD-L1 promoter binding, thereby enhancing PD-L1 expression[35,36].
2.1.3 The HIF-1α pathway: PD-L1 regulation in the hypoxic microenvironment
HIF-1α is a master regulator of cellular adaptation to hypoxia and is widely activated in solid tumors[37,38]. In the hypoxic TME, HIF-1α not only drives metabolic and angiogenic reprogramming but also contributes to immune evasion.
Specifically, HIF-1α can directly promote PD-L1 transcription and enhance its expression in both tumor and immune cells, establishing a hypoxia-driven immunosuppressive program[39-41]. In addition, HIF-1α regulates angiogenic factors and metabolic pathways that collectively reshape the immune microenvironment, leading to reduced T cell and NK cell activity and facilitating tumor immune escape[42,43]. Hypoxia is also closely associated with therapy resistance and may create a feedback loop that further stabilizes HIF-1α signaling[44,45].
2.1.4 Integrative analysis of shared and context-specific mechanisms
The regulatory networks governing PD-L1 expression exhibit strong context dependency across different TME states. The NF-κB and JAK-STAT pathways function as shared signaling hubs activated by both chemotherapy and RT[13,36]. The cGAS-STING pathway is primarily associated with DNA damage induced by RT, while HIF-1α signaling predominates under hypoxic conditions[30,39,46].
Given this complexity, a single therapeutic strategy is unlikely to effectively overcome PD-L1-mediated immune escape across all tumor contexts. Instead, context-specific intervention strategies targeting NF-κB, JAK-STAT, or HIF-1α signaling may provide more effective immunotherapeutic outcomes[47-50].
2.2 PD-L1 resistance mechanisms based on TME subtypes
Immunophenotyping of the TME represents a significant advancement in recent immuno-oncology research. Immune cell infiltration and stromal characteristics profoundly influence tumor immune escape mechanisms, treatment response, and drug resistance[11]. With the ongoing advancement of tumor immunotherapy, identifying tumors with distinct immune subtypes not only helps predict treatment outcomes but also provides a theoretical basis for designing personalized treatment strategies[51-53].
The distinct characteristics of these immune subtypes influence tumor response to ICIs therapy and the development of resistance mechanisms. In clinical practice, treatment strategies should be tailored to the specific tumor immune subtype. Table 1 below summarizes the immune features, core resistance mechanisms, and ICB response predictions for each TME immune subtype.
Table 1. Summary of immunological characteristics and resistance mechanisms based on TME subtypes.
| TME Phenotype | Main Immune Features | Mechanisms of Drug Resistance | Predicted Response to ICB | Refs |
| Hot/Inflamed | The cells showed high T cell infiltration, elevated PD-L1 expression, and a large number of pro-inflammatory cytokines. | T cell depletion leads to the simultaneous activation of multiple inhibitory checkpoints. | High potential for response, but prone to developing acquired resistance. | [10,11,54,55] |
| Cold/Desert | The cells exhibit low T cell infiltration, low PD-L1 expression, and insufficient antigen presentation capacity. | Failure of antigen priming, insufficient recruitment of effector T cells. | Minimal response, typically exhibits primary resistance. | [10,56-58] |
| Immune-rejecting | Immune cells become trapped around the tumor or within the tumor stroma and are unable to reach the tumor core. | Dense ECM barriers and CAF-induced physical and biochemical isolation. | Weak response; T cells cannot effectively interact with tumor cells. | [10,54,56] |
TME: tumor microenvironment; ICB: immune checkpoint blockade; ECM: extracellular matrix; CAF: cancer-associated fibroblast.
2.2.1 Hot tumors: Adaptive immune resistance
Hot tumors (inflammatory type) typically exhibit high levels of immune cell infiltration and PD-L1 expression, and thus often demonstrate favorable clinical responses during the initial stages of ICIs therapy[11]. However, as treatment continues, metabolic inhibitors (such as lactate and adenosine) accumulate in the TME, gradually hindering immune activity. Research indicates that high concentrations of tumor-derived lactic acid acidify the microenvironment and suppress effector T cell function, thereby correlating closely with resistance to anti-PD-1/PD-L1 therapy[59-61]. Meanwhile, adenosine acts as a key immunosuppressive factor in inflammatory tumors, weakening antitumor immune responses through the A2A/A2B receptor signaling pathway. This may also counteract the fundamental immune advantage of hot tumors[62].
2.2.2 Cold tumors: Multidimensional immunosuppression
In contrast, “cold tumors” are typically in a state of immune silencing or an “immune desert”, meaning that immune cells rarely penetrate the tumor, antigens are not effectively presented, and the immune response is weak[11,63]. This immunosuppressive effect is particularly pronounced in certain types of tumors, including pancreatic cancer and prostate cancer. Studies have shown that restoring the function of DCs is crucial for activating potent anti-tumor immunity. Traditional chemotherapy and radiation therapy promote the release of tumor antigens and the activation of immune cells by inducing immunogenic cell death (ICD) and releasing DAMPs[64-66]. Therefore, enhancing the recruitment and activation of immune cells is considered key to improving the efficacy of immunotherapy for cold tumors.
2.2.3 Immune-excluded tumors: The dual challenge of physical barriers and immunosuppression
Immune-excluded tumors typically present with dense extracellular matrix (ECM) deposition and elevated interstitial fluid pressure (IFP), thereby restricting immune cell and therapeutic drug penetration into the tumor parenchyma, thereby restricting immune cell and therapeutic drug penetration into the tumor parenchyma[67-70]. Among these, cancer-associated fibroblasts (CAFs) are important contributors to this barrier formation; they not only remodel and deposit the ECM but also secrete immunosuppressive factors, inhibit CD8+ T cell infiltration, and promote tumor resistance to ICB therapy[67]. Therefore, for immune-excluded tumors, strategies such as degrading or remodeling the ECM, reducing stromal barriers, modulating abnormal vasculature, blocking TGF-β signaling pathways, and achieving targeted drug delivery can promote T cell re-entry into the tumor parenchyma and thereby improve immunotherapy efficacy[71,72].
2.3 Exosome PD-L1 (Exo-PD-L1)
Recent studies have shown that PD-L1 not only exists on the surface of tumor cells, but can also be released into the bloodstream via exosomes. Exosomes are tiny vesicles secreted by cells that encapsulate a variety of molecules derived from their parent cells, including PD-L1. The PD-L1 on these exosomes (i.e., Exo-PD-L1) is identical to the PD-L1 on the surface of tumor cells and can travel throughout the body via the bloodstream[73,74]. This mechanism enables tumors to exert immunosuppressive effects within the local immune microenvironment, thereby influencing the systemic immune system.
The immune evasion mechanism of exosomal PD-L1 manifests primarily in two aspects: First, Exo-PD-L1 acts as a “decoy” by binding to PD-1 on the surface of peripheral T cells, prematurely inducing T cell exhaustion. This prevents effector immune cells from effectively responding to the tumor[75,76]. Second, the secretion of Exo-PD-L1 diminishes the efficacy of anti-PD-L1 antibodies at the tumor site, as it spreads through systemic circulation to induce systemic immune suppression[77,78].
Research indicates that the level of PD-L1 in exosomes is closely correlated with the degree of tumor immune escape. Consequently, targeted therapy against PD-L1 in exosomes has emerged as a novel direction in current immunotherapy[74]. By downregulating the secretion of Exo-PD-L1 or intercepting its interaction with T cells, it is anticipated that the efficacy of ICIs therapy can be enhanced[79].
2.4 MHC-I downregulation
Antigen presentation is fundamental for T cells to recognize and attack tumor cells, and effective immune killing depends on tumor cells presenting antigens to CD8+ T cells via MHC-I molecules. However, many tumor cells evade T cell recognition by downregulating MHC-I expression through genetic mutations or epigenetic modifications. MHC-I downregulation leads to APD, in which tumor cells cannot effectively display their surface antigens, preventing T cells from recognizing and killing them[80,81].
This process is particularly pronounced in “cold tumors” and certain tumors that inhibit the infiltration of immune cells. In cold tumors, the expression of MHC-I on the surface of tumor cells is significantly reduced, thereby inhibiting the infiltration of T cells into the tumor[82]. For example, epigenetic modifications such as B2M gene mutations or DNA methylation have been shown to be important causes of MHC-I downregulation[83]. Recent experimental data indicate that restoring MHC-I expression can enhance the immune response of tumors and improve the efficacy of immunotherapy[84].
In clinical practice, ICIs, such as anti-PD-1 and PD-L1 antibodies, are important treatment strategies for various tumors. However, in tumors with downregulated MHC I, the efficacy of ICIs is often suppressed because the absence of MHC I leads to impaired antigen presentation and weakens CD8+ T cell-mediated immune responses[85,86]. Based on this mechanism, researchers are working to restore MHC I expression through epigenetic modifications or drug regulation. For example, some small molecule drugs or epigenetic interventions can reverse MHC I downregulation and improve the sensitivity of tumors to immunotherapy[85]. In addition, a strategy of targeting and demethylating MHC I antigen-presenting transcription factors (such as NLRC5) has increased MHC I expression in animal models and enhanced the efficacy of anti-PD-1 therapy, suggesting that this approach has potential clinical application value[87].
3. The Immunomodulatory Role of Chemotherapy in TME Remodeling
Chemotherapy has long been regarded as a cytotoxic strategy that inhibits tumor proliferation[88]. However, accumulating evidence demonstrates that its role extends beyond direct tumor killing, as it can also remodel the TME and modulate host immune responses. In particular, chemotherapy-induced ICD serves as a key mechanism linking tumor cell death to immune activation. However, chemotherapy can simultaneously induce adaptive immune resistance, including PD-L1 upregulation through NF-κB-dependent signaling networks (as discussed in Section 2), thereby complicating the therapeutic synergy between chemotherapy and immunotherapy[89,90].
3.1 Dual immunomodulatory effects of chemotherapy
Although chemotherapy can activate robust anti-tumor immunity through ICD, it also induces compensatory immunosuppressive mechanisms within both tumor and immune compartments[91-93]. These effects include modulation of myeloid cell function, activation of immunosuppressive populations, and upregulation of immune checkpoint molecules, collectively contributing to therapy resistance[94-96]. Table 2 summarizes the immunomodulatory benefits and potential immune negative feedback risks of different chemotherapy drugs.
Table 2. Summary of immunological benefits and potential risks of different types of chemotherapy drugs.
| Chemotherapy Drug | Main Immune Benefits | Potential Risks/Limitations | Refs |
| Platinum-based (Cisplatin, Oxaliplatin) | Induces ICD; upregulates MHC-I expression and enhances antigen presentation. | Compensatory upregulation of PD-L1; notable nephrotoxicity and ototoxicity. | [88,97,98] |
| Anthracyclines (Doxorubicin, Mitoxantrone) | Promotes macrophage polarization toward M1 phenotype; reduces MDSC infiltration. | Commonly causes bone marrow suppression; high doses may suppress peripheral lymphocyte proliferation. | [88,97,99] |
| Low-dose Cyclophosphamide | Selectively depletes Tregs; restores effector T cell activity. | Improper dosing may lead to systemic immunosuppression and leukocyte reduction. | [100-102] |
| Epipodophyllotoxins (Etoposide, Teniposide) | Strong induction of ICD-related factors; promotes DCs maturation. | Cardiotoxicity limits cumulative dose and long-term application. | [103-105] |
ICD: immunogenic cell death; MHC: major histocompatibility complex; MDSC: myeloid-derived suppressor cell; DCs: dendritic cells.
3.1.1 Chemotherapy-induced reprogramming of tumor-associated macrophages (TAMs)
TAMs play a central role in tumor progression, metastasis, and post-treatment relapse[106,107]. TAMs exhibit high plasticity along the M1-M2 polarization axis, where M1-like macrophages are generally tumoricidal, whereas M2-like macrophages promote tumor progression and immunosuppression[108,109]. Within the TME, TAMs predominantly exhibit an M2-like phenotype that supports angiogenesis, immune suppression, and metastatic dissemination[110,111]. Chemotherapy-induced systemic inflammatory responses can further drive TAM polarization toward the M2 phenotype, thereby reinforcing immunosuppressive conditions. NF-κB signaling acts as a context-dependent regulator of immune responses within tumor-infiltrating immune cells[80,112]. It is required for the activation and effector functions of T cells, NK cells, and NKT cells[113], while also sustaining the immunosuppressive activity of regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs)[94]. Importantly, chemotherapy can partially reprogram TAMs toward an M1-like phenotype under specific conditions[114]. However, it may also promote the expansion of immunosuppressive TAM subsets (e.g., CXCL7+ TAMs), which contribute to chemoresistance through CXCL7-CXCR2 signaling and downstream tumor-promoting pathways[115].
3.1.2 Reactive oxygen species (ROS)
ROS play a context-dependent dual role in chemotherapy-induced tumor immune regulation[116,117]. Moderate ROS levels promote endoplasmic reticulum stress and the release of DAMPs (calreticulin, adenosine triphosphate (ATP), high mobility group box 1 (HMGB1)), thereby enhancing DC activation and T cell priming through ICD[118]. Ferroptosis-associated lipid peroxidation can further amplify this immunogenic effect[119,120]. In contrast, sustained oxidative stress is associated with immunosuppression and immune escape. Increased ROS levels correlate with upregulation of PD-L1 in tumor and immune cells, contributing to adaptive resistance[121,122]. Chemotherapy-induced oxidative stress, such as that triggered by paclitaxel (PTX) or gemcitabine, has been linked to transient PD-L1 elevation in tumor cells and macrophages[121]. Overall, the effects of ROS are highly dose- and duration-dependent, promoting either ICD-mediated immune activation or immune suppression depending on stress intensity[123-125].
3.1.3 Positive effect of inducing ICD
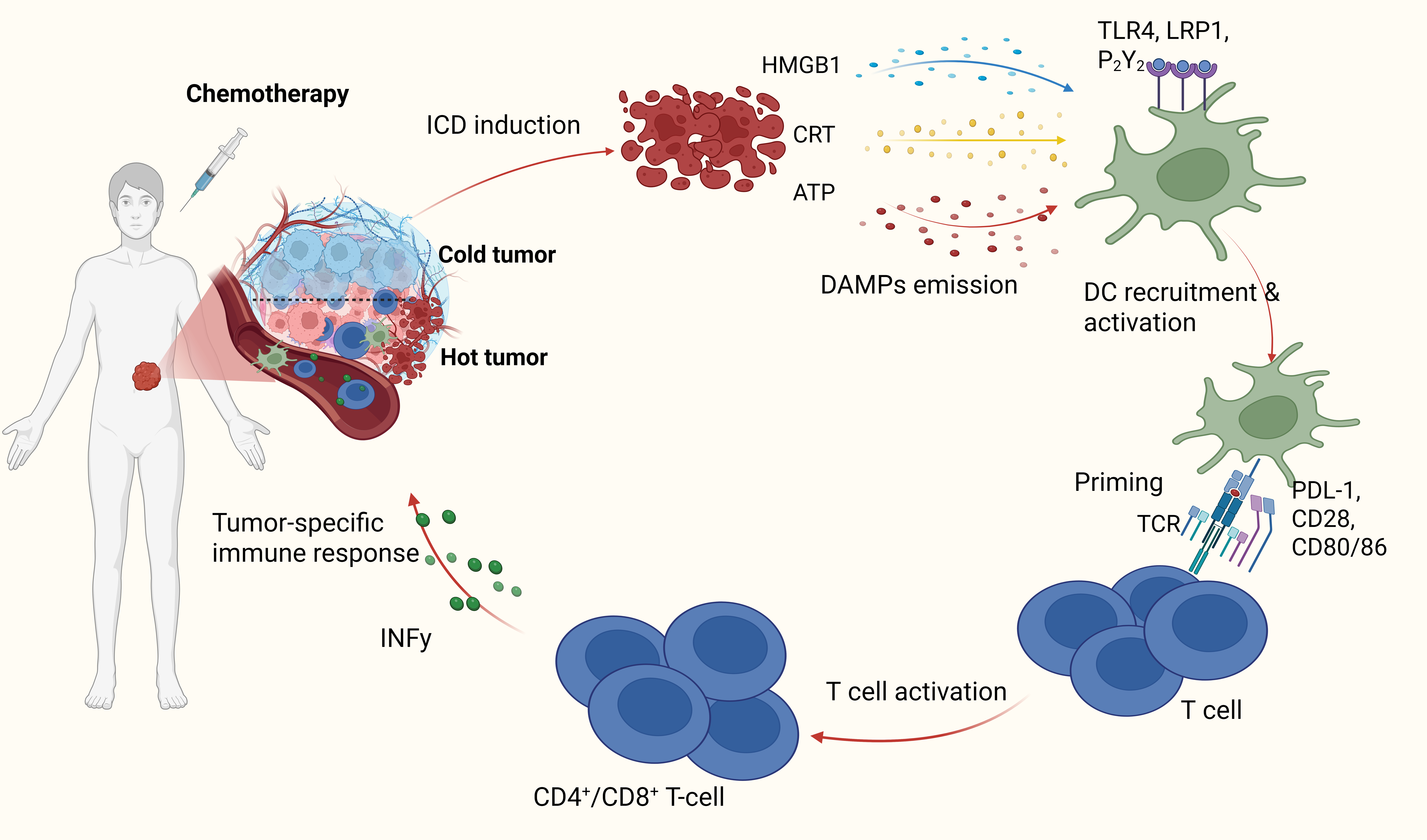
ICD is a regulated form of cell death characterized by the emission of DAMPs, which convert dying tumor cells into endogenous vaccines that activate anti-tumor immunity[11,126-129]. Chemotherapeutic agents such as cisplatin, oxaliplatin, and PTX can induce ICD by triggering cellular stress responses, thereby promoting the transition of the TME from an immune-cold to an immune-hot state (Figure 3).
{kind=link}
Figure 3. Illustration of chemotherapy-induced ICD and its role in converting cold tumors into hot tumors. Created in BioRender. Lutf, B. (2026) https://BioRender.com/93z6ui0. ICD: immunogenic cell death; INF: interferon; DC: dendritic cell; TCR: T-cell receptor; TLR: toll-like receptor; LRP: low-density lipoprotein receptor-related protein; HMGB: high mobility group box; CRT: calreticulin; ATP: adenosine triphosphate; DAMPs: damage-associated molecular patterns.
During ICD, CRT exposure acts as an “eat-me” signal that promotes DC recognition and phagocytosis of tumor antigens. Subsequently, HMGB1 released from dying cells interacts with pattern recognition receptors such as TLR4 to enhance antigen-presenting cell (APC) maturation and cytokine production[129-131]. In parallel, extracellular ATP functions as a chemotactic and inflammasome-activating signal that promotes T cell priming and expansion[132,133].
Activated DCs further stimulate CD4+ and CD8+ T cell responses through co-stimulatory signaling pathways such as CD28-CD80/86[134,135]. Although PD-1/PD-L1 signaling can modulate this process, ICD overall enhances tumor antigen presentation and T cell clonal expansion, thereby improving the efficacy of subsequent immunotherapy[136].
3.2 Optimization of chemotherapy efficacy under nano-intervention
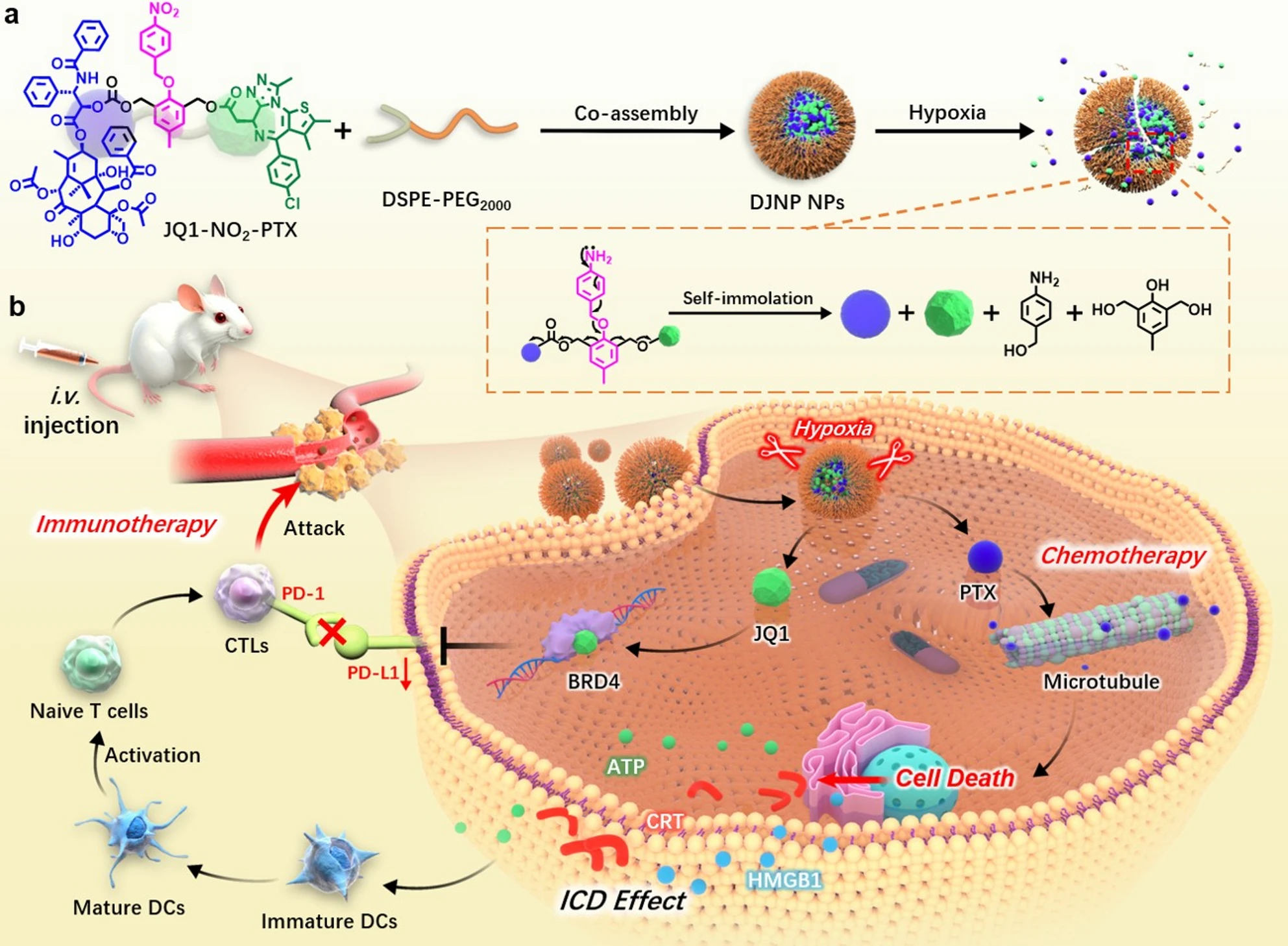
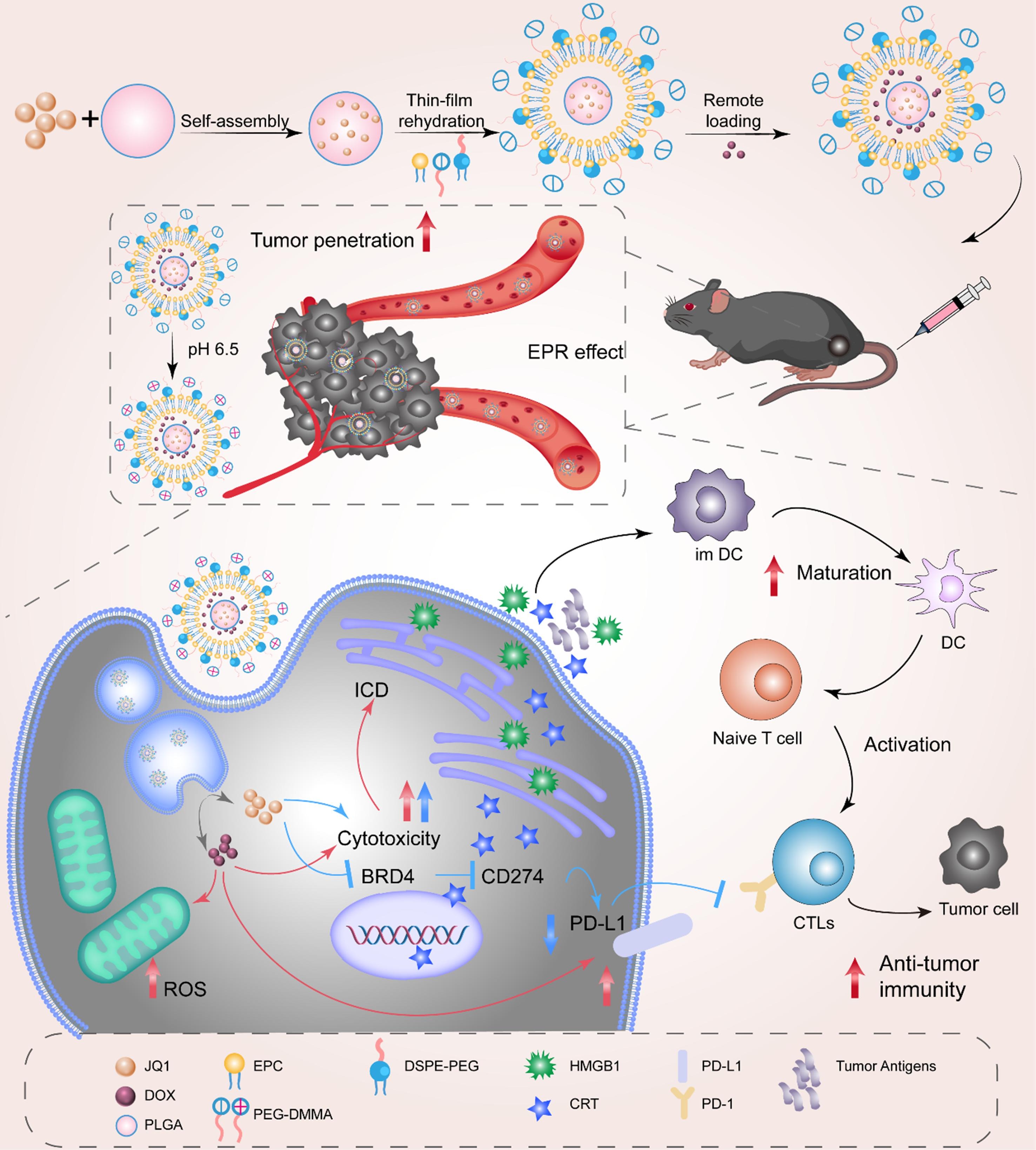
To overcome the adverse effects of traditional chemotherapy in immunotherapy, particularly immune resistance caused by upregulation of the PD-L1/PD-1 axis, nanotechnology emerges as a novel therapeutic strategy, providing critical support for enhancing chemotherapy efficacy while reducing toxicity[137-139]. To overcome this problem, researchers designed and developed a hypoxia-responsive nanoprodrug (DJNP nanoparticles), which combines PTX with the BRD4 inhibitor JQ1, and utilizes the hypoxia microenvironment to trigger the activation of the prodrug, precisely releasing the therapeutic drug (Figure 4)[140]. JQ1, as a BRD4 inhibitor, reduces PD-L1 expression by inhibiting the binding of BRD4 to the PD-L1 promoter. Studies have shown that DJNP nanoparticles can reverse chemotherapy-induced PD-L1 upregulation, effectively enhancing the tumor’s immune response by reducing PD-L1 expression and blocking tumor cell immune escape. In mouse models, DJNP nanoparticles exhibited higher tumor accumulation and prolonged blood circulation time, and compared to the traditional chemotherapy drug Abraxane, they more effectively inhibited tumor growth without showing significant systemic toxicity. In another study, Hu et al. constructed a polydopamine nanocarrier that could synergistically load doxorubicin (DOX) and the BRD4 inhibitor JQ1, enabling JQ1 to inhibit PD-L1 expression at the molecular level and significantly enhance the anti-tumor synergistic effect in vivo[141]. Similarly, Liu et al. designed a hormone-reversing eggshell liposome system that co-loaded the PD-L1 inhibitor JQ1 and the chemotherapeutic drug DOX, and enhanced the chemotherapeutic immunotherapy effect on tumors through this nanoplatform (Figure 5)[142]. These eggshell liposomes exhibited good drug loading stability in the TME and achieved drug release in the acidic TME through a pH-responsive mechanism. JQ1 inhibited PD-L1 expression, thereby blocking the tumor immune escape pathway; simultaneously, DOX-induced ICD further enhanced the anti-tumor immune response. In vivo experiments showed that JQ1/DOX eggshell liposomes not only significantly inhibited tumor growth but also enhanced the immune system’s recognition and clearance of tumors by increasing CD8+ T cell infiltration and inhibiting PD-L1 expression.
{kind=link}
Figure 4. (A) Schematic illustration of the preparation of hypoxia-responsive self-immolative DJNP NPs; (B) Their application in chemo-immunotherapy via PTX-induced ICD and JQ1-mediated PD-L1 downregulation. Reproduced from reference[140]. CC BY 4.0. DJNP NPs: prodrug nanoparticles; PTX: paclitaxel; ICD: immunogenic cell death; DCs: dendritic cells; BRD4: bromodomain-containing protein 4; DSPE: 1,2-distearoyl-sn-glycero-3-phosphoethanolamine; PEG: polyethylene glycol.
{kind=link}
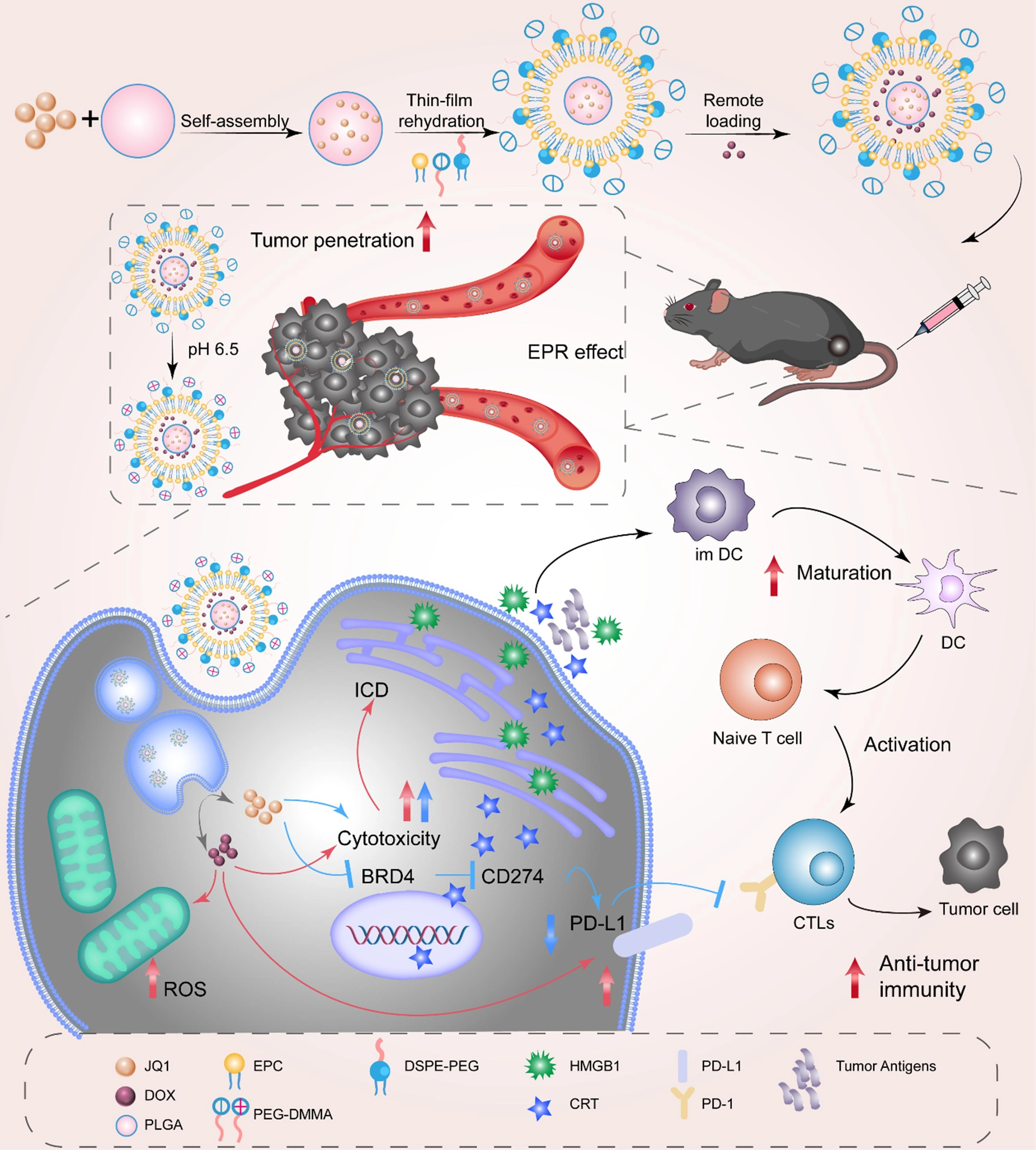
Figure 5. The illustration shows how charge-reversed eggshell liposomes can be loaded with DOX and JQ1 to improve chemotherapy-immunotherapy by blocking the PD-L1 pathway. Reproduced with permission from reference[142]. Copyright © 2023 Elsevier. DOX: doxorubicin; ICD: immunogenic cell death; BRD4: bromodomain-containing protein 4; ROS: reactive oxygen species; DC: dendritic cell; CTLs: cytotoxic T lymphocytes; PLGA: poly(lactic-co-glycolic acid); PEG: polyethylene glycol; HMGB: high mobility group box; CRT: calreticulin; DSPE: 1,2-distearoyl-sn-glycero-3-phosphoethanolamine.
4. Local Effects of RT
RT, as a traditional local treatment, has long primarily reduced tumor volume through physical means. However, with the rapid development of tumor immunology, the role of RT has gradually transcended simple tumor control, becoming an important “igniter” in tumor immunotherapy[143,144]. RT can not only directly kill tumor cells, but more importantly, it can reshape the TME, transforming physical damage into strong immune activation signals, thereby stimulating the host’s immune response, overcoming the tumor’s immune escape mechanism, and effectively reversing the state of immune resistance[32].
4.1 RT-induced immune activation and adaptive resistance
RT exerts potent immunomodulatory effects by inducing DNA DSBs, which represent the primary trigger of radiation-driven immune activation (see Section 2.1.2 for detailed cGAS-STING signaling mechanisms)[145]. These DNA damage events ultimately convert irradiated tumors into highly immunogenic sites, promoting anti-tumor immune responses[143,146].
Importantly, RT-induced immune activation is tightly regulated by dose and temporal dynamics. While moderate radiation doses enhance immune priming, excessive irradiation may attenuate immune stimulation through negative feedback mechanisms, highlighting the importance of optimized fractionation strategies[143,147-149].
However, RT simultaneously induces adaptive immune resistance. Radiation-triggered interferon signaling can promote upregulation of immune checkpoint molecules such as PD-L1 in both tumor and immune cells[20]. This dynamic feedback represents a key limiting factor in sustained anti-tumor immunity and underscores the importance of timing in RT-immunotherapy combinations.
4.2 Application of nanotechnology in radiosensitization and immunomodulation
Traditional RT still faces several challenges, such as severe side effects and limited local control rates due to insufficient absorption of radiation by cancerous tissue and exposure to high-dose ionizing radiation, which ultimately affect patient prognosis. Therefore, it is crucial to avoid residual cancer cells, as these cells may develop RT resistance. It has been reported that pretreatment with low-dose radiation (LDR) before RT can significantly reduce tumor growth and invasiveness[150].
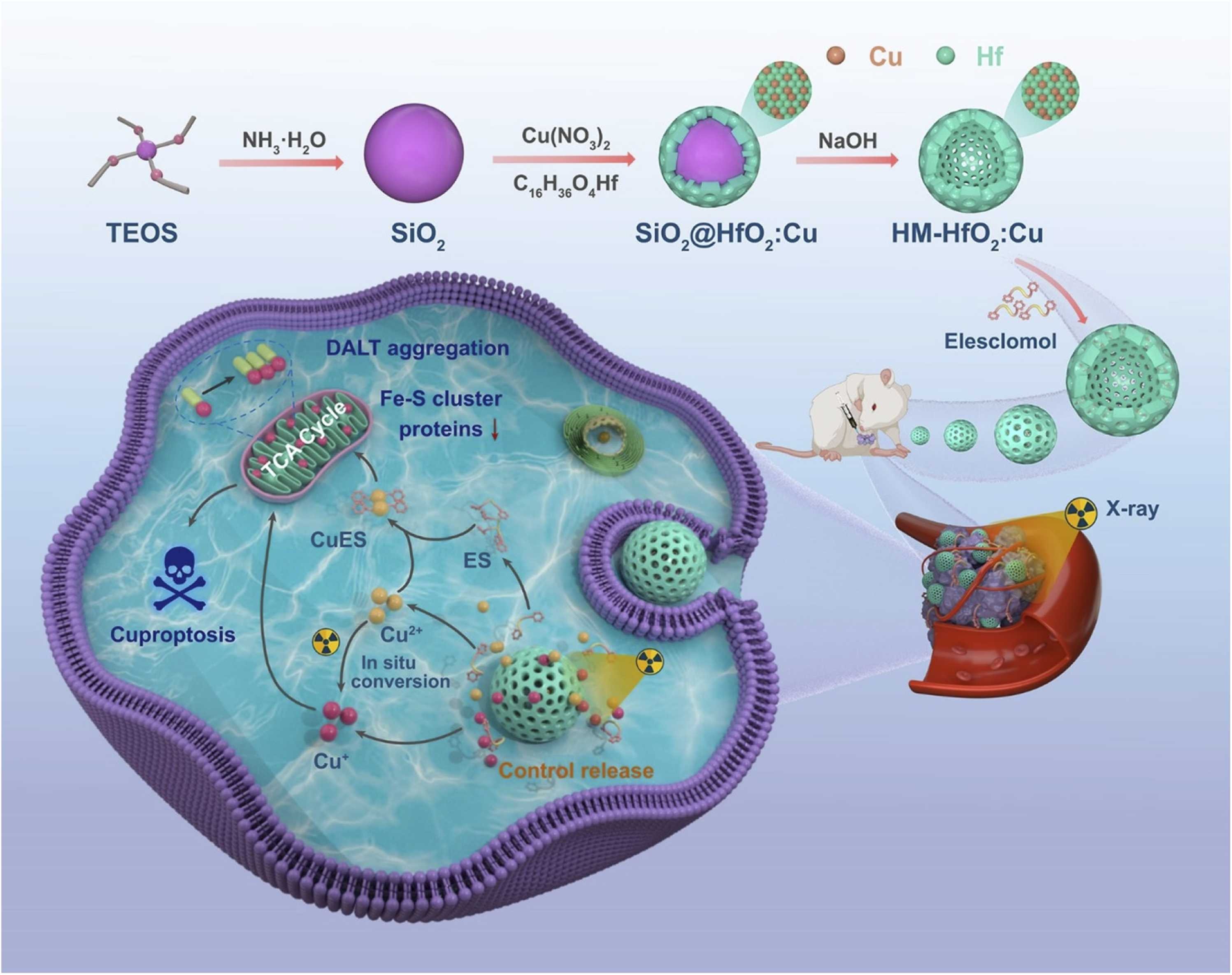
Advances in nanotechnology have significantly broadened the application of RT in cancer treatment. To enhance the effectiveness of RT, radiosensitizers are widely used in the process[151,152]. Radiation therapy using High-Z nanoparticles as radiosensitizers is a key method for increasing the radiation dose delivered to the target area during treatment. Gold (Au), hafnium (Hf), and bismuth (Bi) are examples of high-atomic-number elements that absorb X-rays very well. When these substances accumulate in tumor tissue in the form of nanoparticles, they enhance the photoelectric effect, generating large numbers of secondary electrons and Auger electrons. This exacerbates localized DNA damage and makes tumor cells more sensitive to radiation therapy[150,153]. For example, hafnium oxide nanoparticles NBTXR3 based on high-Z elements not only significantly enhance the local efficacy of RT but also promote immune cell infiltration at tumor sites and induce abscopal effects in distant tumors. This demonstrates that high-Z nanoscale sensitization technology can also promote RT-associated antitumor immune responses[154]. Recently, a copper-doped HfO2 nanocage was developed as an RT sensitizer (Figure 6)[155]. In this system, Cu ions are doped into the HfO2 nanocapsule shell, while elesclomol, a copper ionophore, is loaded into the hollow mesoporous structure. Upon X-ray irradiation, the nanocapsules release Cu ions, which may undergo intracellular redox conversion between Cu(II) and Cu(I). Since Cu(I) is generally considered the bioactive copper species associated with cuproptosis, the released copper can promote mitochondrial lipoylated protein aggregation and iron-sulfur cluster protein loss, thereby enhancing tumor cell death and radiosensitization.
{kind=link}
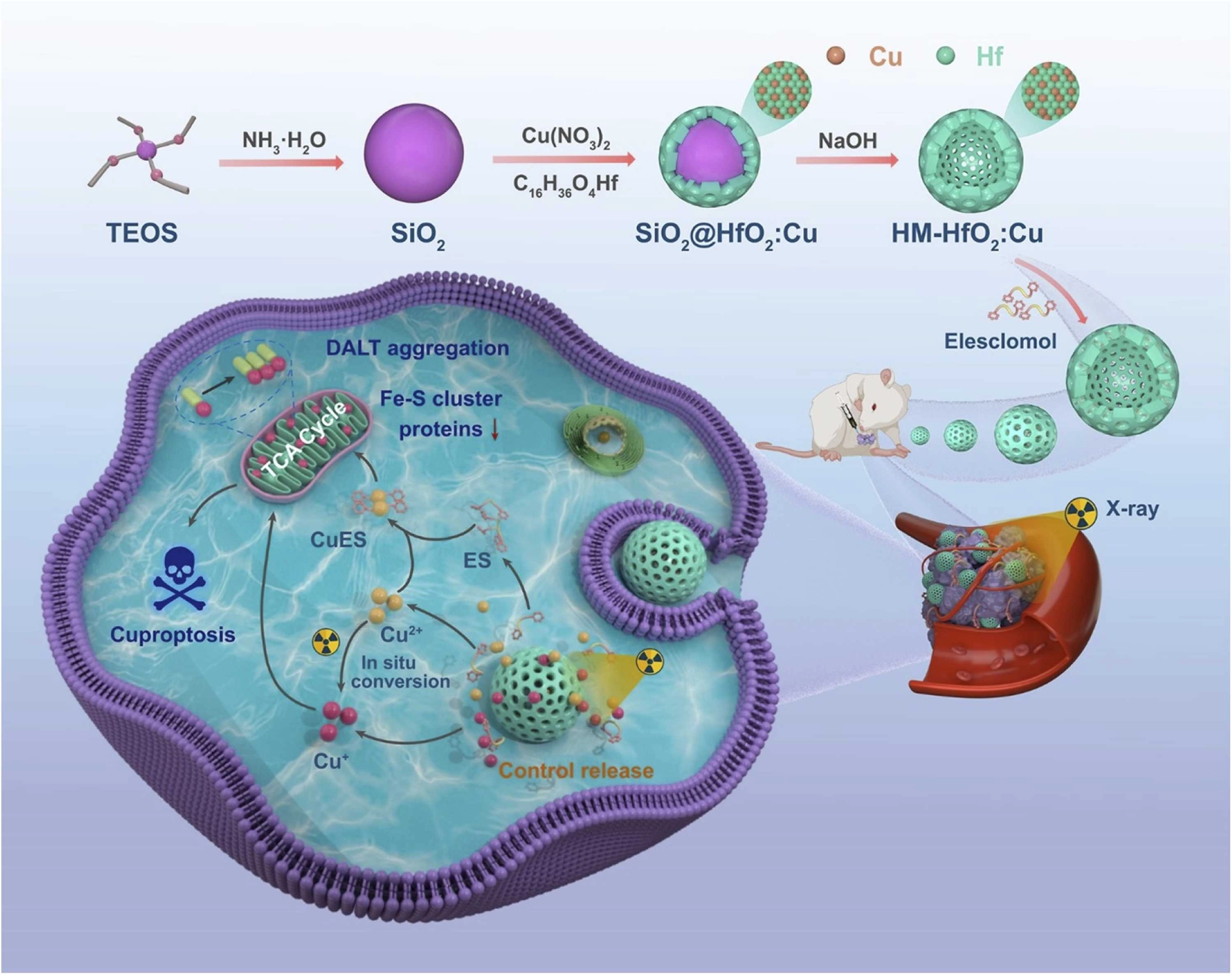
Figure 6. Schematic illustration of ES@HM-HfO2 tumor radiosensitization. Upon X-ray irradiation, the HfO2-based nanocapsules release and reduce Cu ions, which are transported to mitochondria by encapsulated ES. The accumulated Cu+ disrupts Fe-S cluster proteins and promotes DLAT aggregation, triggering cuproptosis and enhancing radiotherapeutic efficacy. Reproduced with permission from reference[155]. Copyright © 2025 Elsevier. ES: elesclomol; HM: hollow mesoporous; DLAT: dihydrolipoamide S-acetyltransferase; TEOS: tetraethyl orthosilicate.
In nanotechnology applications, triphenylphosphine-derived metformin (Tpp-Met)@MnO2@Alb nanoparticles demonstrate significant potential as a novel radiosensitizer for enhancing the efficacy of tumor RT (Figure 7)[156]. Research indicates that Tpp-Met@MnO2@Alb enhances RT efficacy through a dual mechanism: On one hand, it inhibits oxidative phosphorylation in tumor cells via Tpp-Met, effectively reversing the tumor’s hypoxic environment and amplifying DNA damage induced by RT; on the second hand, it improves the tumor immune microenvironment by suppressing PD-L1 and TGF-β1 expression, thereby overcoming post-RT immune tolerance. Specifically, this therapeutic strategy enhances tumor-specific immune responses by activating the cGAS-STING pathway. This pathway stimulates immune cells by sensing tumor-derived dsDNA induced by RT, further promoting DC maturation and activating T cells, thereby enhancing antitumor immunity. Therefore, Tpp-Met@MnO2@Alb nanoparticles not only enhance the efficacy of localized RT but also activate RT-induced antitumor immune responses through immunomodulation. Specifically, by activating the cGAS-STING signaling pathway, they provide more persistent and extensive immune memory. As nanotechnology continues to advance in cancer treatment, it holds promise for playing a greater role in enhancing RT sensitization and immunomodulation in the future.
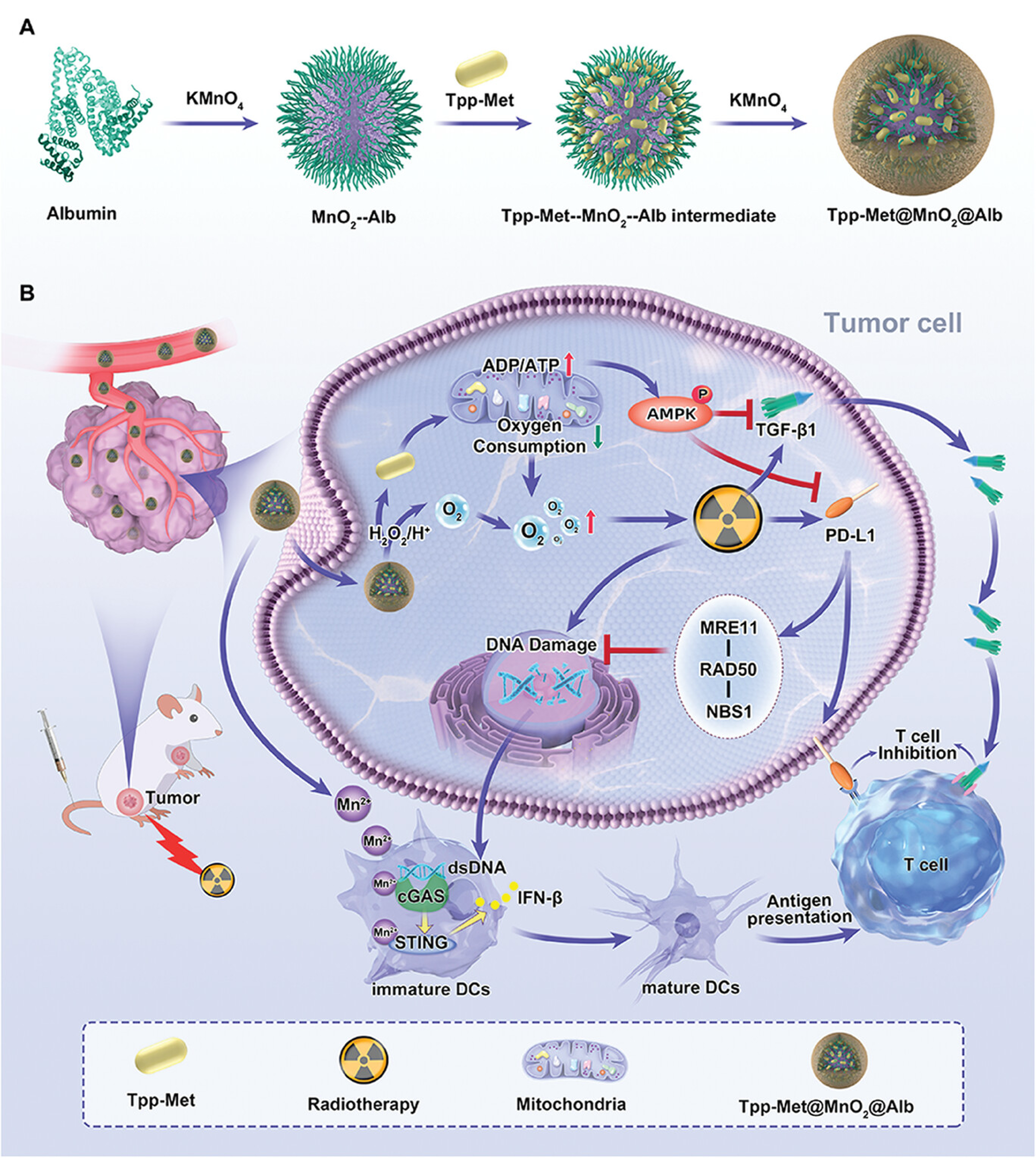{kind=link}
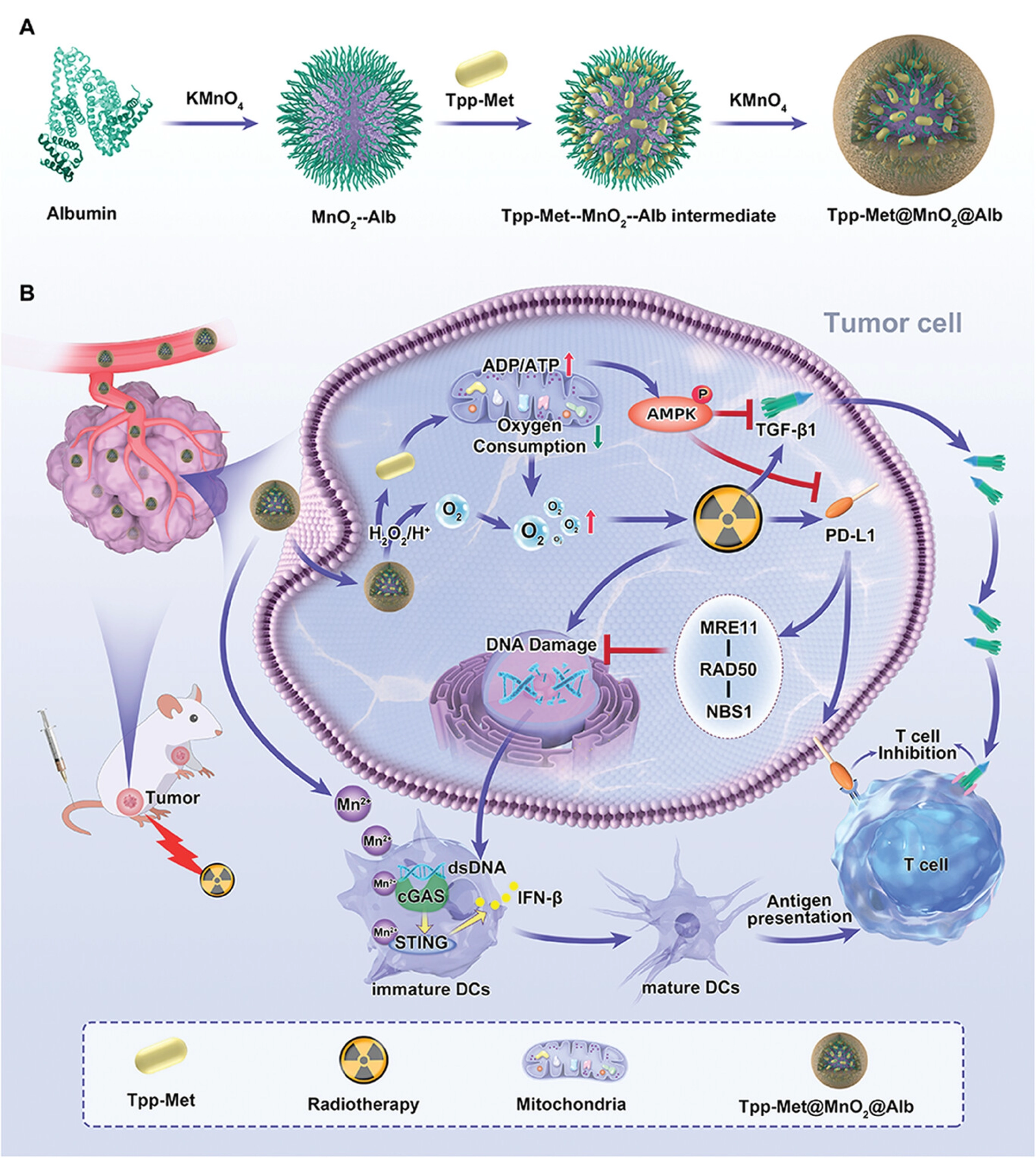
Figure 7. Schematic illustration of Tpp-Met@MnO2@Alb nanoparticles for enhanced radio-immunotherapy. The nanoparticles were prepared by biomineralization and improve therapeutic efficacy through PD-L1 and TGF-β1 downregulation, cGAS-STING pathway activation, and two-stage oxygen modulation. Reproduced with permission from reference[156]. Copyright © 2024 John Wiley & Sons. TGF: transforming growth factor; cGAS: cyclic GMP-AMP synthase; STING: stimulator of interferon genes; ADP: adenosine diphosphate; ATP: adenosine triphosphate; AMPK: AMP-activated protein kinase; dsDNA: double-stranded DNA; DCs: dendritic cells.
5. The Mechanism of Synergy Between Chemotherapy and Radiation Therapy
The therapeutic synergy between chemotherapy and RT extends beyond direct cytotoxicity and is closely associated with the restoration of anti-tumor immune surveillance. This process is characterized by the release of immunogenic signals, including CRT exposure, extracellular ATP, and HMGB1 release, which collectively promote DC activation and adaptive immune priming[157]. In parallel, RT-induced DNA damage activates the cGAS-STING pathway, serving as a central hub that integrates genotoxic stress with type I interferon signaling, thereby enhancing anti-tumor immunity and immune surveillance[158]. Importantly, the combination of chemotherapy and RT amplifies tumor immunogenicity and creates a more favorable immune microenvironment for immunotherapy. However, this synergy is often limited by systemic toxicity and suboptimal drug delivery, highlighting the need for advanced therapeutic strategies.
5.1 Synergistic application of nanotechnology in chemotherapy and RT
As mentioned earlier, chemotherapy is a commonly used and effective treatment method in clinical practice, while CRT is an organic combination of chemotherapy and RT. Combined CRT is more effective at controlling local tumors than single therapy because it works synergistically. However, it still has many problems, such as the inability to deliver enough drugs to the tumor site and the serious side effects on healthy tissues[151,159,160]. Nanotechnology plays a crucial role in the combined application of CRT and immunotherapy. Researchers can utilize multifunctional nanoplatforms that not only deliver drugs and immunomodulatory molecules to tumor sites with high precision, ensuring their proper delivery, but also leverage the unique physicochemical properties of nanomaterials to significantly enhance therapeutic synergy, thereby overcoming the limitations of conventional therapies[161-163].
Platinum-based drugs are not only widely used in chemotherapy, but also serve as highly effective radiosensitizers[151,164]. For example, lactose-modified self-targeting nanoprodrugs utilize ASGPR-mediated endocytosis to precisely enrich hepatocellular carcinoma cells, and the released cisplatin significantly enhances radiosensitivity by inducing DNA damage and arresting cells in the G2/M phase (Figure 8)[165]. Similarly, the AgNPs@TEOS-MTX platform developed by Soroushnasab et al. utilizes MTX-mediated active targeting to achieve highly efficient drug enrichment in glioblastoma (GBM). This system combines the radiosensitizing effect of silver nanoparticles with the cytotoxicity of MTX, further enhancing the synergistic therapeutic effect by inducing large-scale apoptosis[166].
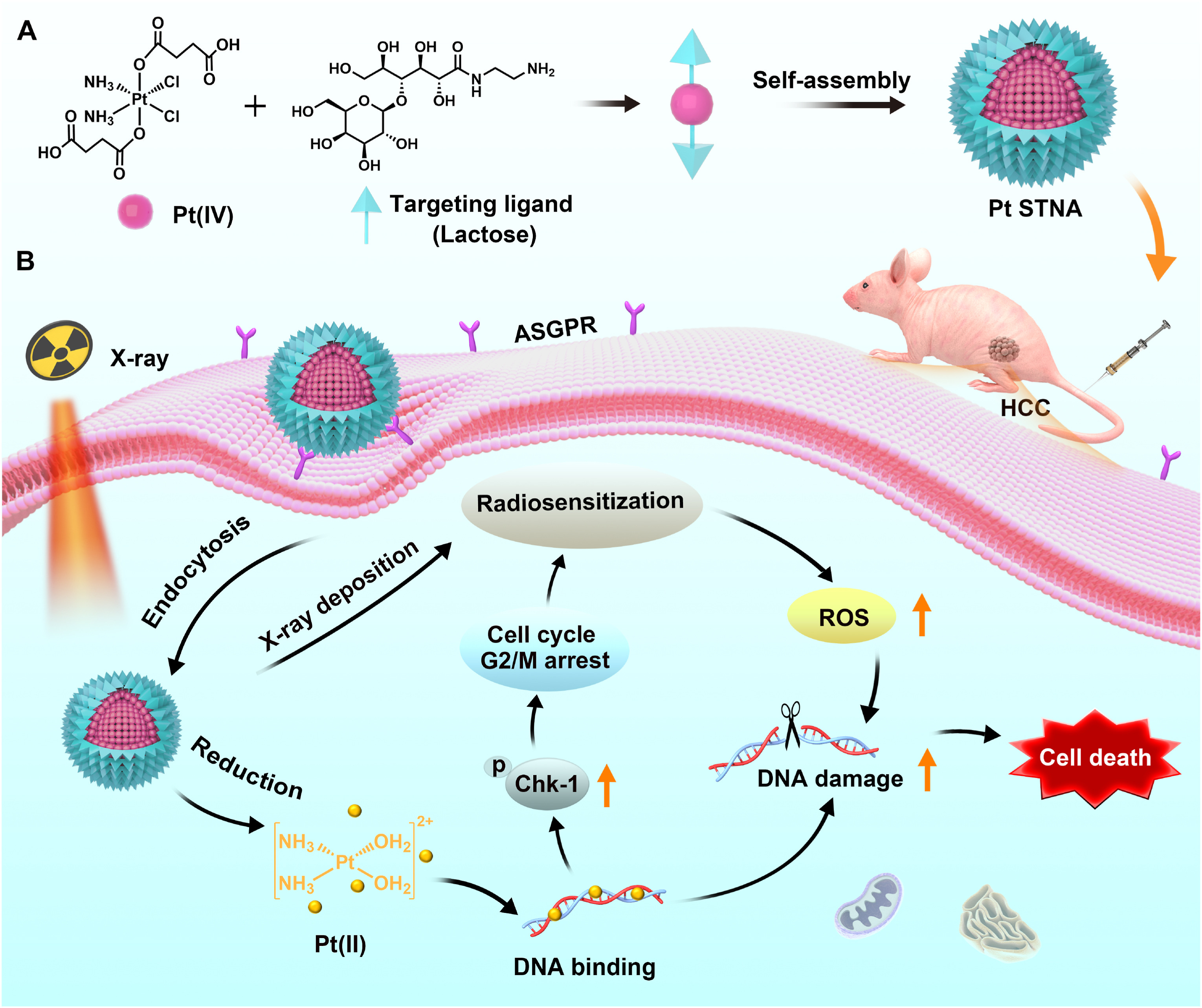{kind=link}
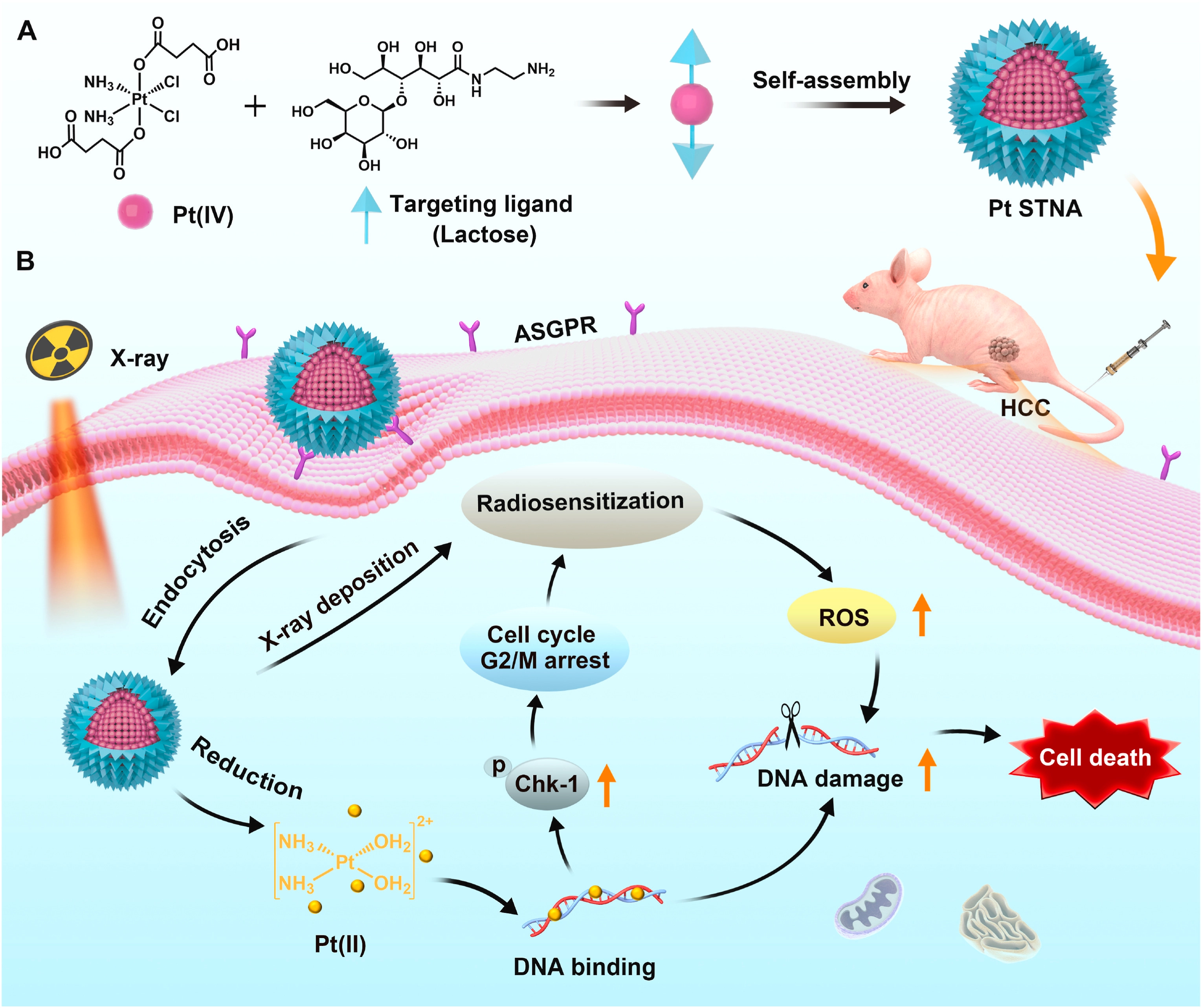
Figure 8. Schematic illustration of platinum (IV) amphiphilic Pt STNA and its radiosensitizing mechanism for synergistic CRT of hepatocellular carcinoma. Reproduced with permission from reference[165]. Copyright © 2022 Elsevier. Pt STNA: prodrug-based self-targeting nanoassembly; CRT: chemoradiotherapy; ASGPR: asialoglycoprotein receptor; ROS: reactive oxygen species; HCC: hepatocellular carcinoma.
In addition, in response to the challenges of blood-brain barrier blockage and significant side effects in the treatment of GBM, another study developed an X-ray activated nanoprodrug platform (Figure 9)[167]. This system uses downconversion nanoparticles as the core, coated with X-ray sensitive materials, which can release chemotherapeutic drugs in situ under LDR excitation. This design not only realizes imaging-guided CRT synergy, but also allows researchers to assess the dynamic changes in the microvascular environment in real time through visualization, thereby providing precise morphological evidence for clinical decision-making. This shift from “blind treatment” to “visualized diagnosis and treatment” represents an important direction of evolution in nanomedicine.
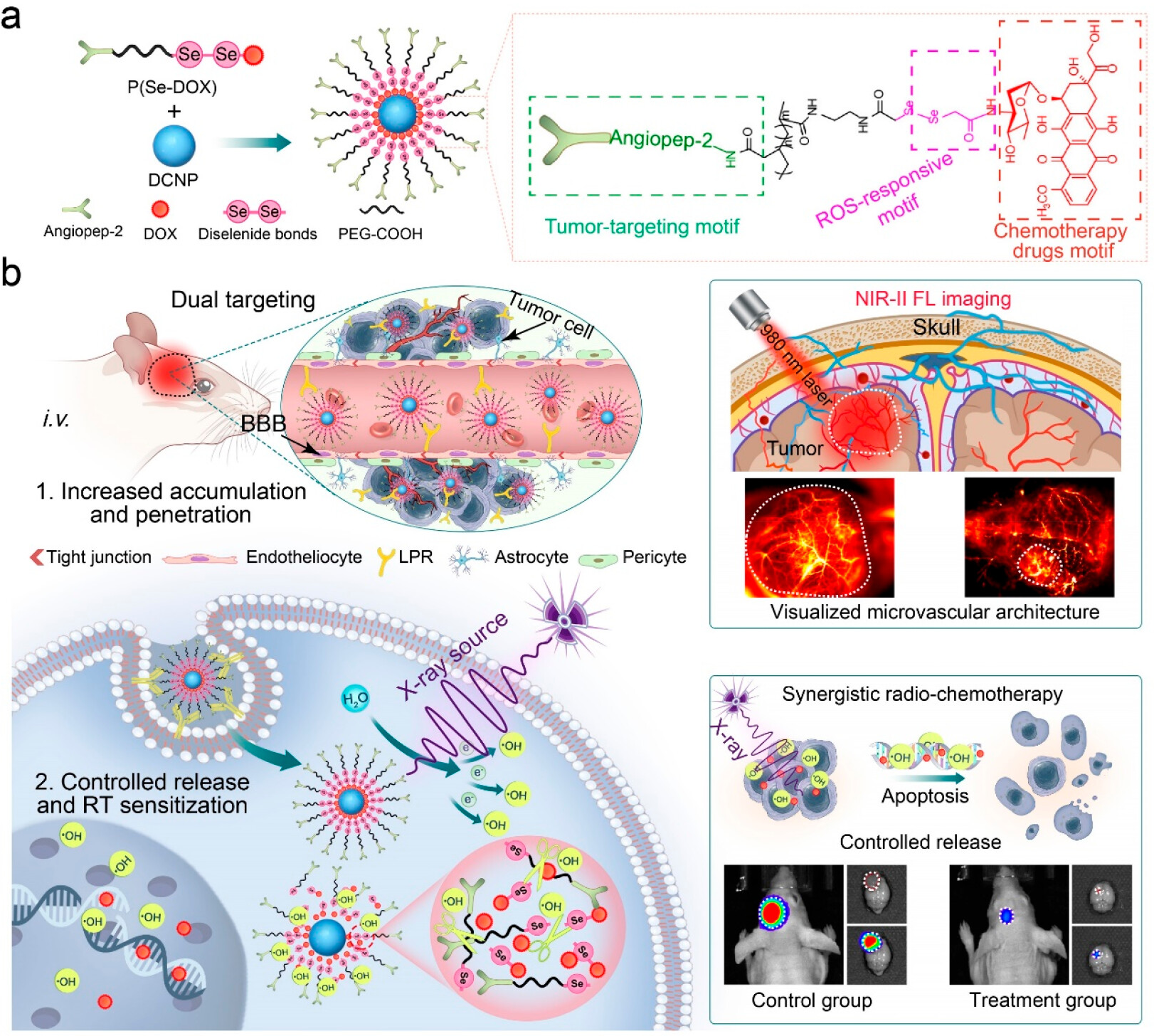{kind=link}
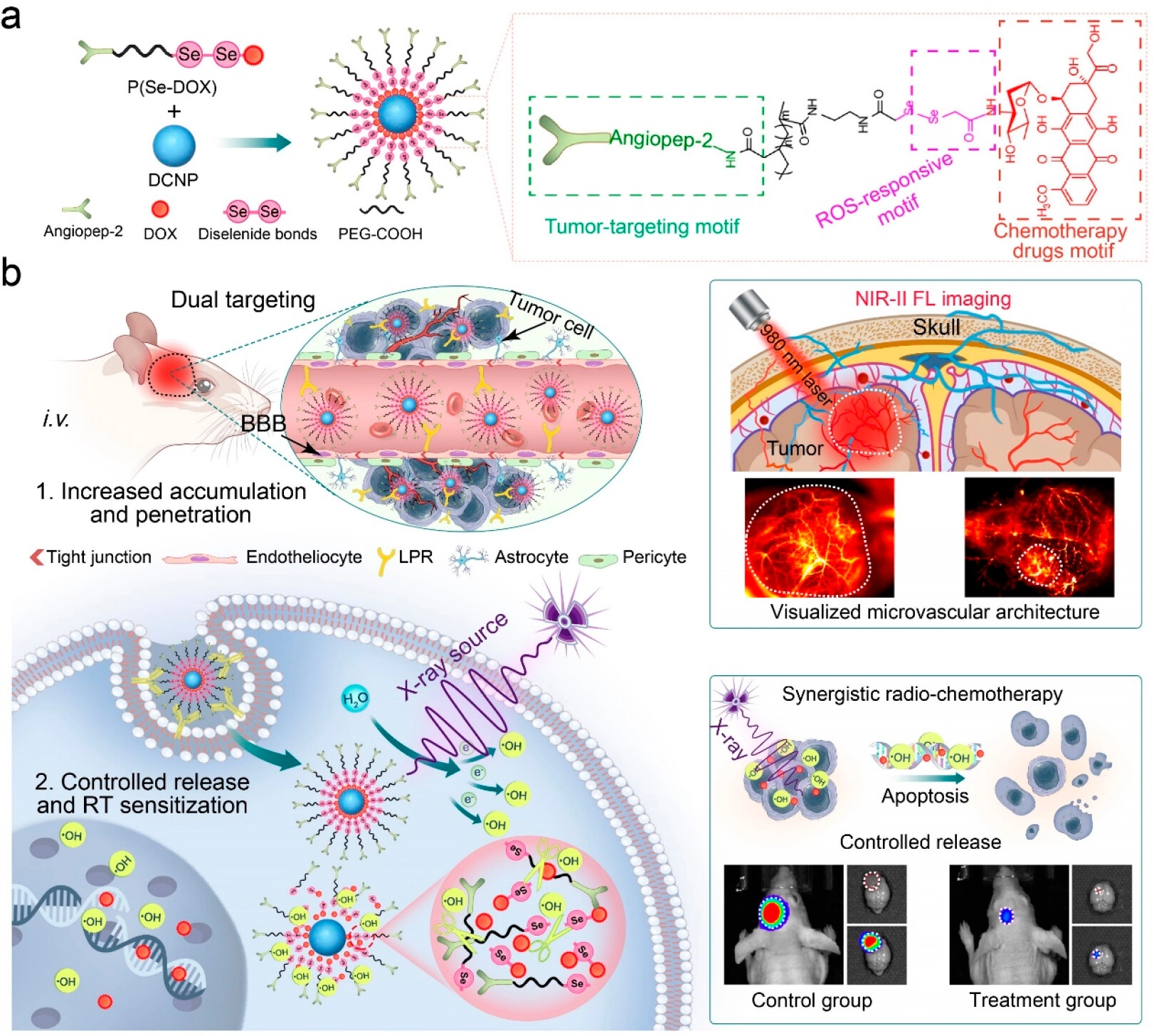
Figure 9. DCNP@P(Se-DOX)@ANG performs thetargeted penetration of BBB, and NIR-II FL imaging maps the microvascular structure of glioma and guides the mechanism and application of synergistic antiglioma radiation-chemotherapy. Reproduced with permission from reference[167]. Copyright © 2024 American Chemical Society. BBB: blood-brain barrier; NIR: near-infrared; FL: fluorescence; DOX: doxorubicin; DCNP: downconversion nanoparticle; PEG: polyethylene glycol; ROS: reactive oxygen species; RT: radiotherapy.
6. Finding the Optimal Intervention Window for Time-Series Strategies
In this section, “sequential nanoregulation” refers to the coordinated control of treatment timing, therapeutic sequence, and stimulus-responsive drug release. It integrates three levels of regulation: (1) temporal control of CRT and immunotherapy administration, (2) sequential drug release enabled by nanocarriers, and (3) combination strategies that align biological response windows with therapeutic intervention.
6.1 Dynamic evolution of PD-L1 expression and its time-dependent characteristics
PD-L1/PD-1 expression appears to follow a dynamic temporal pattern that may influence the synergy between ICIs and chemotherapy. Multiple studies suggest that a 3-5 days window may represent a favorable period for combining immunotherapy with chemotherapy in certain settings - though this conclusion warrants a stratified interpretation: (1) In chemotherapy-dominant regimens, particularly among lung cancer patients, the evidence supports a 3-5 day window. Yao et al. found that administering anti-PD-1 therapy on day 3 after chemotherapy (e.g., cisplatin) was associated with reduced chemotherapy-induced lymphocyte toxicity, and its clinical efficacy was superior to concurrent or first-line immunotherapy. A retrospective analysis of patients with advanced lung cancer further confirmed that administration of ICIs within 1-10 days after chemotherapy (especially days 3-5) was associated with significantly improved overall survival (OS) compared to other time arrangements (17.3 months vs. 12.7 months)[168]. From an immunological perspective, Huo et al.’s research revealed the biological basis of this phenomenon: the third day after chemotherapy is a period of increased or peak-like PD-1 expression on CD8+ T cells. Clinical data show that, compared with concurrent treatment, this 3-day delayed sequential treatment regimen significantly improved the objective response rate (68% vs. 37%) and disease control rate (98% vs. 81%), while effectively prolonging progression-free survival (PFS)[169]. (2) In the case of RT alone or combined CRT, the relevant evidence is highly heterogeneous, and the timing of ICI administration after RT appears to be more variable and may extend beyond the early post-treatment phase depending on experimental or clinical conditions. Similar to the rapid induction induced by chemotherapy, RT-induced PD-L1 expression exhibits a significant “hysteresis effect”, indicating a critical therapeutic window. Studies have shown that the 3rd to 5th day after RT is a favorable time to introduce ICIs. Lee et al. demonstrated that injecting anti-PD-L1 antibodies on day 5 after mice received a 10 Gy radiation dose most effectively inhibited tumor growth, because at this time, the activity of cytotoxic T lymphocytes (CTLs) in the tumor is closely related to the peak expression of PD-L1, and precise blocking time can interrupt the inhibitory signal, thereby maintaining the killing ability of CTLs[170]. Yi et al. used dynamic SPECT/CT imaging to identify a critical window period of PD-L1 upregulation following RT, demonstrating that anti-PD-1 intervention during this window significantly enhanced anti-tumor immune responses[171].
However, in complex clinical practice, the process of tumor immune microenvironment remodeling induced by CRT exhibits more dynamic and individualized characteristics. Pouw et al. used whole-body positron emission tomography imaging to track ICI uptake in NSCLC patients during CRT and came to an unexpected conclusion: in the first week after treatment, local tumor drug uptake did not increase as expected, but remained stable or even decreased, while uptake in peripheral immune organs such as the bone marrow and spleen fluctuated significantly[172]. This finding highlights that extrapolating from a single point in time may not be applicable to all clinical situations. Similarly, Zhong et al. combining immune cell and PD-L1 imaging techniques, found that in some combined CRT regimens, the optimal window for immunotherapy can be extended to day 14 after RT, at which point intervention produces the most significant immune enhancement effect[173].
In addition to macroscopic regulation based on the CRT induction cycle, the “circadian rhythm of infusion” at the microscopic level has gradually become an important dimension affecting the timing of intervention. Landré et al. conducted a meta-analysis on the impact of immunotherapy infusion time on the survival of patients with advanced cancer[174]. Specifically, patients who received early infusion before 13:00 (including NSCLC, melanoma, etc.) had significantly better OS and PFS than those who received late infusion, with hazard ratios as low as 0.50 and 0.51, respectively. This suggests that early infusion may optimize the rhythmic function of immune cells and produce a better synergistic effect with the immunomodulatory effect of PD-L1, further enhancing the overall efficacy of immunotherapy. This suggests that early infusion may optimize the rhythmic function of immune cells and produce a better synergistic effect with the immunomodulatory effect of PD-L1, further enhancing the overall efficacy of immunotherapy. This finding may be explained by circadian regulation of immune cell trafficking and activation, as immune cell proliferation, cytokine secretion, and T cell responsiveness exhibit time-of-day-dependent fluctuations governed by systemic circadian clocks.
Overall, the current evidence suggests that the optimal immunotherapy intervention window is not a fixed time point but rather a dynamic and context-dependent period shaped by tumor type, treatment modality, dosing schedule, and host immune status. Therefore, dynamic imaging technology for monitoring systemic immune patterns has irreplaceable value in determining personalized dosing regimens and optimizing clinical treatment strategies.
6.2 Nanocarrier platforms for spatiotemporally controlled drug delivery
Due to the significant spatiotemporal mismatch between chemotherapy/RT and immunotherapy in inducing antitumor immune responses (Section 6.1), traditional systemic combination therapies struggle to precisely capture the optimal intervention window. Nanotechnology, through meticulous functionalization design, provides an ideal carrier platform for the clinical translation of sequential strategies[139,152,175].
The multiresponsive nature of nanocarriers enables sequential drug release, allowing precise coordination between chemotherapy-induced immune activation and the desensitization process of immunotherapy. For instance, Wang et al. developed a pH/reduction dual-responsive self-assembling nanoparticle that first releases DOX in the acidic TME to induce ICD, followed by the release of immunostimulants within DCs. The sequential release mechanism of “ignition first, acceleration later” greatly improves the response rate of tumor-infiltrating CD8+ T cells[176]. Similarly, Reda et al. successfully delivered PLK1 inhibitors and PD-L1 antibodies simultaneously using a mesoporous silica nanomaterial system (Antigen Release Agent and Checkpoint inhibitor). This strategy not only enhanced the immune response to lung cancer but also significantly reduced systemic toxicity, fully demonstrating the unique advantages of nanocarriers in sequential synergistic therapy[177].
In addition to endogenous environmental responses, external physical stimuli can also enhance the dynamic regulation capabilities of drug delivery systems. Studies have shown that polymer nanosystems responsive to NIR-II (second near-infrared window) can immediately induce a strong ICD effect upon photoexcitation, thereby promoting the controlled release of siPD-L1 and the reconstruction of the immune microenvironment[178]. This light-controlled release method has high spatiotemporal resolution, making it easy to pinpoint the exact time and location for immune intervention. In summary, carrier systems utilizing nanomaterials can simultaneously deliver CRT and immunotherapy, and the drug release time can be controlled very precisely through responsive structural design. This “on-demand release” characteristic breaks through the inherent time limitations of traditional drug delivery, thus helping to achieve precise timing of immune interventions[179].
7. Multimodal Embedding and Spatiotemporal Controlled Release in Nanotechnology
Given the complexity and heterogeneity of the TME, single treatment methods are often insufficient to effectively overcome PD-L1 resistance. Therefore, the development of precise and personalized therapeutic strategies is of paramount importance. By combining multi-target nanocarriers with PD-L1 inhibitors and adding them to chemotherapy, RT and other treatments, immune responses can be significantly improved[139,180]. Recent studies have further demonstrated that nanosystems can enhance tumor immunotherapy through multiple coordinated interactions rather than a single delivery function. For example, pH-responsive and cascade-responsive nanoplatforms have been designed to integrate tumor targeting, antigen capture, PD-L1 silencing, DC modulation, and T cell reprogramming within a single therapeutic system. These findings suggest that multifunctional nanosystems may improve antitumor immunity by simultaneously regulating tumor cells, APCs, and the immunosuppressive microenvironment[181,182]. Table 3 summarizes the comprehensive applications and therapeutic effects of various nanomaterials in a variety of diseases. This section will explore various approaches to treating different types of tumors that affect the immune system, with a focus on real-time treatment options for “cold tumors”, “hot tumors”, “immune-rejecting tumors”, and tumors that cannot effectively present antigens. Furthermore, it will investigate intelligent controlled release technology, the role of PD-L1 in exosomes, and methods for dynamically evaluating the TME.
Table 3. The comprehensive applications and therapeutic effects of various nanomaterials in a variety of diseases.
| Cancer types | Nanomaterials | Nanoplatform type | Co-treatment Strategy | Mechanisms of Immune Evasion | Therapeutic Effects | Limitations (Cons) | Refs |
| 4T1 & MB49 | IR-LND@Lip | Lipid-based nanocarrier | RT | Co-inhibition of PD-L1, collagen, and TGF-β to overcome immune escape in solid tumors. | Significantly enhanced sensitivity to RT by reversing tumor microenvironment, boosting immune activation, and suppressing tumor growth and metastasis. | Liposomes have limited long-term storage stability; lipid-cell membrane interactions may lead to off-target effects. | [183] |
| LLC | SAB-NO NPs | Polymeric nanoparticles | RT | Reverses tumor hypoxia, co-inhibits PD-L1 and COX-2 to overcome immune suppression, enhances T-cell infiltration. | Sensitizes RT, induces ICD, enhances T-cell activation and infiltration, and suppresses tumor growth. | NO release kinetics may be variable; there may be systemic vasodilatory side effects; and precise control of NO release in vivo is quite difficult. | [184] |
| B16F10 | NPs-MΦ | Biomimetic nanoparticles | Adoptive macrophage therapy | CD133+ PD-L1+ cancer cells upregulate PD-L1 and TGF-β signaling, causing ECM stiffening and resistance to macrophage phagocytosis. | TGF-β inhibition enhances macrophage-mediated tumor cell killing, improves tumor suppression, and overcomes PD-L1 resistance. | Macrophages are complex to source and engineer; batch-to-batch consistency of cell function is difficult to guarantee; large-scale production presents significant challenges. | [185] |
| B16F10 | GELN | Plant-derived nanoparticles | a-PD-L1 | Modulation of gut microbiota to enhance unsaturated fatty acid (DHA) production, reducing PD-L1 expression in tumors. | GELN enhances anti-PD-L1 efficacy by inducing DHA production, inhibiting PD-L1 expression, and reducing tumor growth and metastasis. | Plant-derived materials exhibit significant batch-to-batch compositional variability; the mechanism relies on gut microbiota regulation, with significant individual differences; and long-term safety data are limited. | [186] |
| B16-OVA & E.G7-OVA | NanoST | Phytoglycogen nanoparticles | Intradermal vaccination-PD-1 blockade | Tumors express PD-L1, inhibiting CD8+ T cell activation. NanoST enhances DC activation and CD8+ T cell response. | NanoST induced CD8+ T cell activation, significantly reduced tumor growth, and when combined with PD-1 blockade, provided strong antitumor immunity. | The effects of immune adjuvants may vary depending on individual immune status; optimal combinations with other ICIs require further optimization. | [187] |
| Various cancers | MOFs | Metal-organic frameworks | - | Tumors exhibit immune escape via PD-L1 expression; TME alters tumor response to therapy. | MOFs enhance RT effects and anti-PD-L1 response via spatiotemporal control, improving immune activation and overcoming resistance. | There are uncertainties regarding in vivo stability and the biosafety of degradation products; controlling structural uniformity in large-scale production is difficult; and the regulatory approval pathway is unclear. | [188] |
| 4T1 | MUC-1 aptamer-conjugated Ag/MSN Janus nanoparticles | Janus nanoparticles | Chemo-PDT-Gene therapy | TME acidity promotes drug release; MUC-1 overexpression leads to immune evasion. | pH-responsive release system improves therapeutic efficiency; synergistic effect of chemotherapy, photothermal therapy, and gene therapy. | Multifunctional design increases manufacturing complexity; potential cytotoxicity of silver nanoparticles; batch consistency of Janus structure is difficult to control. | [189] |
| GBM | poly(MIs)/PTX@PEI/siPGK1@CCM | Biomimetic polymer nanoparticles | CRT | Hypoxic TME induces therapy resistance; PGK1 overexpression enhances tumor energy metabolism and survival | Enhanced tumor suppression via PGK1 silencing, improved PTX sensitization and RT response, extended survival in orthotopic GBM models | Quality control of cancer cell membrane-derived siRNA is complex; large-scale production of multilayer structures (polymer + siRNA + cell membrane) presents challenges; batch-to-batch variation in siRNA stability and delivery efficiency. | [190] |
| B16-F10 & 4T1 | D/UCNPs, cgAuNCs | Upconversion nanoparticles/Gold nanoclusters | Chemo-PTT-NIR-II imaging | Nanoparticles responsive to TME acidity/oxidation; release therapeutic agents and generate ROS→enhanced ICD | Enhanced tumor imaging; synergistic chemo-PDT efficacy; increased ROS, ICD, improved immune activation and tumor suppression | The long-term in vivo accumulation and toxicity of rare earth elements; the upconversion efficiency is limited by tissue penetration depth; the long-term metabolic pathways of gold nanoclusters remain to be elucidated. | [191] |
| 4T1 & LLC | PD-L1 siRNA-BNCT | Boron nanoparticles | RT-Immunotherapy | PD-L1-mediated immune suppression; TME immunosuppressive signaling | Reduced PD-L1 expression, enhanced radiosensitivity, improved T cell activation, stronger tumor suppression | Boron neutron capture therapy requires specialized facilities; boron has limited enrichment efficiency in tumors; and siRNA delivery stability needs optimization. | [192] |
| B16-F10 | Anti-PD-1 siRNA-encapsulated NPs | Liposomal nanoparticles | PD-1 blockade | PD-1-mediated immune suppression, TME immune evasion | Enhanced T-cell mediated immune response, tumor growth inhibition, improved survival | Issues include the in vivo stability and immunogenicity of siRNA; strict storage requirements for liposomes; and the limited duration of gene silencing efficiency. | [193] |
| B16-F10 | aPD-1NCs & aOX40 | ROS-responsive nanogels | Immunotherapy | TME ROS response, PD-1-mediated immune suppression; OX40 agonist enhances T cell co-stimulation, PD-1 inhibition releases immune suppression | Increased T cell activation, immune response, and tumor suppression, enhanced therapeutic efficacy through ROS-responsive release | Heterogeneity in intratumoral ROS levels affects the accuracy of responsive release; the biosafety of gel degradation products needs to be assessed; and the risk of immune-related adverse events associated with dual immune agonists is also considered. | [194] |
| CT26 | ICB-LPs | Liposomes | ICB | PD-L1 recycling and immune suppression; multivalent binding promotes lysosomal PD-L1 degradation and T cell activation | Promoted durable PD-L1 degradation, enhanced CD8+ T-cell responses, and potent antitumor immunity in tumor-bearing mice | Optimization of liposome stability and circulating half-life is required; multivalent binding may increase the risk of off-target binding | [195] |
| 4T1 | piLNP | Lipid nanoparticles | PTT - anti-PD-L1 | Tumor immune escape via PD-L1; immunosuppressive TME limits immune cell activity | Effective tumor targeting, enhanced PTT efficacy, activation of antigen-specific T cells, prevention of metastasis/recurrence | Photothermal therapy is limited by tissue penetration depth; the thermal stability of lipid nanoparticles needs to be considered; and the therapeutic effect on deep tumors may be limited. | [196] |
| 4T1 | CuET@PHF | Polymer-based nanoparticles | PTT and Immunotherapy | Hypoxic TME suppresses cuproptosis proteins; TME normalization enhances cell death and immunogenic responses | Suppressed tumor growth, eliminated CSCs, reduced recurrence/metastasis, enhanced tumor immune responses through TME regulation | Systemic toxicity risks of copper ions; tumor specificity of the copper cuproptosis mechanism needs to be validated; long-term safety needs to be evaluated. | [197] |
RT: radiotherapy; TGF: transforming growth factor; TME: tumor microenvironment; NO: nitric oxide; NPs: nanoparticles; ICD: immunogenic cell death; ECM: extracellular matrix; DHA: docosahexaenoic acid; GELN: ginger-derived exosome-like nanoparticles; OVA: ovalbumin; DC: dendritic cell; MOFs: metal-organic frameworks; MSN: mesoporous silica nanoparticles; PDT: photodynamic therapy; GBM: glioblastoma; PTX: paclitaxel; PEI: polyethylenimine; CCM: cancer cell membrane; CRT: chemoradiotherapy; UCNPs: upconversion nanoparticles; PTT: photothermal therapy; NIR: near-infrared; ROS: reactive oxygen species; BNCT: boron neutron capture therapy; siRNA: small interfering RNA; ICB: immune checkpoint blockade; LPs: liposomes; PHF: PEGylated hollow framework; CSCs: cancer stem cells.
7.1 Treatment decisions based on TME subtyping
7.1.1 Cold tumors typically
This type of tumor typically presents with insufficient infiltration of immune cells and impaired antigen presentation mechanisms[198,199]. To address the problem of “cold” tumors, scientists have created a pH-responsive peptide-based nanorobot (NRb)[200]. It can recognize and bind to PD-L1 on the surface of colorectal cancer cells. In an acidic TME (pH 6.5), NRb transforms from nanoparticles into nanofibers, mechanically disrupting cell membranes and inducing ICD, promoting the release of DAMPs (ATP, HMGB1, CRT) and activating DCs, thereby significantly enhancing CD3+, CD4+, and CD8+ T cell infiltration in PMMR colorectal cancer. Simultaneously, the formed fibrous network can sustainably block the PD-1/PD-L1 pathway for over 120 hours, achieving a synergistic effect of antigen release and ICB.
In addition, other researchers have proposed a TPP-TAM@Alb nanosystem[201]. This system enhances the enrichment of PTX@Alb in tumors and improves the ICD effect by activating the AMP-activated protein kinase (AMPK) pathway to inhibit TGF-β expression and reduce collagen deposition, while inhibiting PD-L1 upregulation and promoting the transition from a “cold” microenvironment to a “hot” microenvironment. Additionally, Sun et al. employed nano-delivered PDLIM2 expression plasmids (nanoPDLIM2) in a mouse lung cancer model. When combined with PD-1 blockade and chemotherapy drugs, this approach significantly increased MHC-I expression, enhanced T-cell infiltration, and successfully converted cold tumors into hot tumors. This strategy markedly amplified the synergistic effects of PD-1 blockade combined with chemotherapy. In most mice, this treatment regimen even achieved complete tumor regression[202].
7.1.2 Immune-exclusion tumors typically
Manifesting as immune cells becoming trapped within the tumor’s peripheral stroma, these tumors are unable to effectively penetrate into the tumor’s core region[203-205]. The convective pressure generated by high IFP significantly restricts the migration of nanoparticles across blood vessels[206]. To overcome this problem, nanotechnology has achieved a systematic reshaping of the tumor biophysical environment through a “physical dismantling” effect. Theoretical simulations and case studies have confirmed that nano-intervention-induced tumor vascular normalization is key to restoring the intravascular and extravascular pressure gradient. The mathematical model developed by Sirin Yonucu et al. demonstrates that by pruning dysfunctional dilated microvessels, nanoscale systems can drive a fundamental shift in drug delivery patterns, transforming the traditional peripheral accumulation around tumors of micelles, nanoprobes, or liposomes into deep penetration into the tumor core[207]. Zhou et al. used the IR-TAM nanoplatform to target mitochondria and activate the AMPK pathway, which not only improved hypoxia but also reduced PD-L1 and TGF-β expression, thereby reducing ECM collagen deposition and enhancing T cell infiltration. Simultaneously, it improved radiosensitivity and inhibited radiation-induced pulmonary fibrosis (Figure 10)[208]. Further studies confirmed that combined blockade of PD-L1 and TGF-β not only downregulates immunosuppressive factors by activating the AMPK pathway but also reshapes the T cell landscape at the cellular differentiation level. Specifically, it induces the expansion of stem-like CD8+ T cells and promotes their differentiation into highly effective effector cells. Concurrently, CAFs maintain the immunosuppressive microenvironment of immune-rejecting tumors by secreting factors like TGF-β. The combined blockade of PD-L1/TGF-β reverses this suppression. Coupled with the IFN-γ-triggered “interferon-licensing” effect across the entire microenvironment, this significantly enhances T-cell infiltration dynamics. Ultimately, immune-rejecting tumors successfully transition to an inflammatory phenotype, markedly improving clinical outcomes[209].
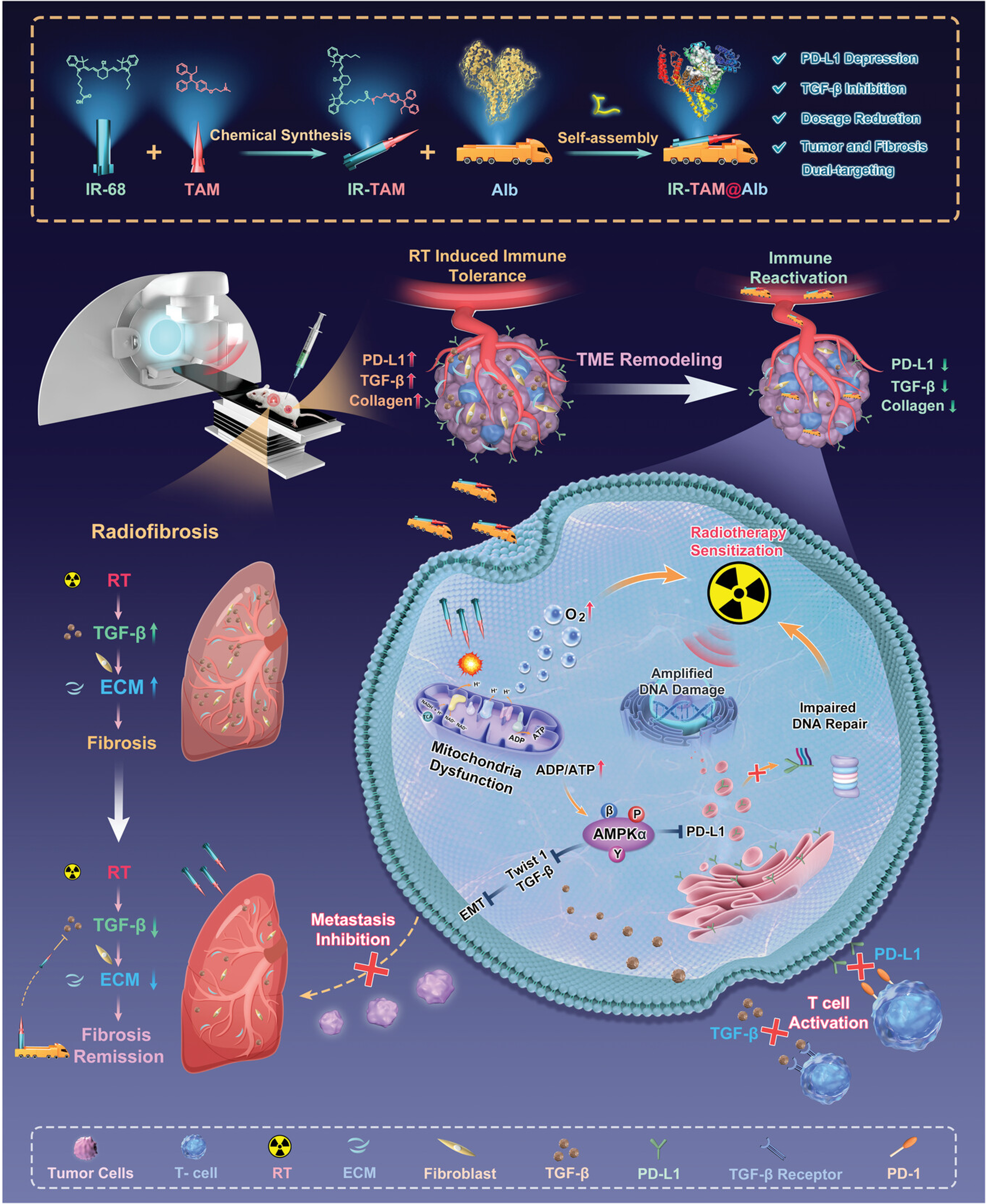{kind=link}
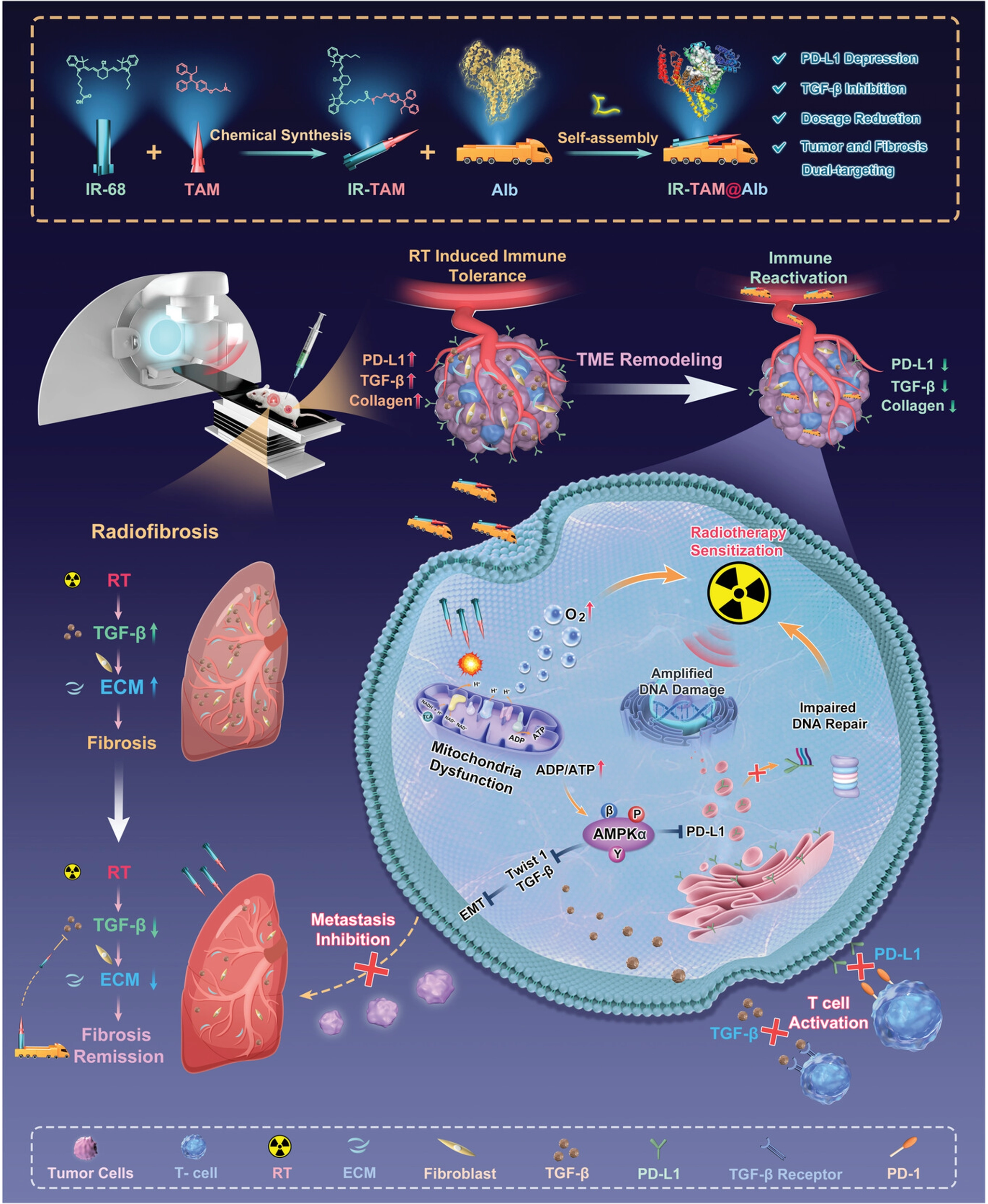
Figure 10. The Preparation Process of IR-TAM@Alb Nanoparticles and Mechanism for Enhancing Radioimmunotherapy and Attenuating RIPF Through Simultaneous Targeting of TGF-β and PD-L1. Reproduced from reference[193]. CC BY 4.0. RIPF: radiation-induced lung fibrosis.
7.1.3 Hot tumors
Thermal tumors exhibit high levels of immune cell infiltration, but are affected by PD-L1 and Exo-PD-L1-mediated immunosuppression[11]. The apatinib-ExoaPD-L1 exosome platform enhances targeting through aPD-L1 modification and synergistically induces ICD and promotes DC maturation with apatinib, while reducing the proportion of Tregs and MDSCs, thereby enhancing CD8+ T cell infiltration and improving anti-tumor immune responses[210]. Furthermore, the IR-FFA@Alb nanosystem activates the AMPK/mTOR pathway to simultaneously downregulate PD-L1 and CD276, and regulates lipid metabolism (promoting FAO and inhibiting glycolysis) to reduce lactate levels, thereby enhancing CD4+ and CD8+ T cell infiltration, promoting long-term immune memory formation, and reducing the risk of tumor recurrence[211].
7.2 Cellular remodeling
To overcome the immunosuppressive TME, nanotechnology-enabled strategies have been developed to reprogram immune cells and restore anti-tumor immune activity.
For TAMs, biomimetic nanoplatforms provide an effective approach to promote M1-like polarization and enhance immune activation. For example, Wang et al. used natural signaling molecules on the membrane surface to encapsulate PLGA nanoparticles within the membrane of M1 macrophages, thereby accurately reprogramming the phenotype of TAMs[212]. Experimental results show that reprogrammed M1 macrophages can reactivate exhausted CD8+ T cells by releasing IL-12 and TNF-α. This can inhibit the growth of primary breast tumors and reduce the likelihood of distant metastasis. Besides directly delivering agonists, changing the pathways that process intracellular miRNA has become a new strategy. Zhang et al. found that targeting and upregulating Dicer expression can effectively reverse the TAM phenotype in colorectal cancer liver metastases[213]. Activating Dicer promotes the processing and maturation of tumor-suppressing miRNAs and blocks M2 polarization signals. This triggers a robust antitumor immune response and prolongs survival. To further enhance precision and reduce systemic side effects, bioorthogonal chemistry has been introduced into microenvironment remodeling. Zhang et al. employed gold nano-enzymes to catalyze the in situ release of “locked” toll-like receptor (TLR 7/8) agonists within macrophages. This in situ triggering mechanism not only enabled polarized directional induction but also significantly enhanced macrophage phagocytic activity toward tumors, offering a novel approach to overcome the toxic side effects of traditional immune agonists[214]. Additionally, the nanoplatform demonstrates exceptional multidimensional synergistic potential. For instance, the combination of TLR 7/8 agonist-loaded nanovaccines with sunitinib and PD-L1 inhibitors, where sunitinib reduces infiltration to optimize the metabolic microenvironment while ICB unleashes T-cell efficacy, successfully elicited durable antitumor immune memory[215].
7.3 Smart controlled-release
As described in Section 3.1.2, ROS exhibit a dose-dependent role in tumor immune regulation. Taking advantage of the unique TME, nanoplatforms can utilize elevated ROS levels and acidic pH as endogenous triggers to achieve spatiotemporally controlled drug release. These stimuli-responsive systems enable sequential and programmed delivery of therapeutic agents within the TME, thereby providing a material basis for combination and staged therapeutic strategies. For example, a study developed a pH/ROS dual-responsive autocatalytic release system that enables deep-tissue delivery of immunotherapeutic drugs under acidic and high ROS conditions, significantly enhancing immunotherapy efficacy and prolonging survival in tumor model mice[216]. Another study engineered a pH/ROS dual-responsive co-delivery nanosystem for liver cancer therapy, achieving synergistic controlled release under TME conditions. This demonstrates that responsive controlled release facilitates the implementation of combination treatment strategies[217]. Additionally, by modulating the pH and redox responsiveness of nanocarriers, ROS generation and immune activity can be enhanced, thereby boosting the synergistic antitumor effects of photodynamic therapy and immunotherapy[218]. Furthermore, researchers have engineered a dual ROS-responsive nanogel platform capable of sequentially releasing immune agonists (e.g., OX40 agonists) to activate T cells, followed by ICIs (e.g., PD-L1 antibodies) to block tumor immune escape. This achieves a precise sequential strategy of “antigen release - immune activation - blockade intervention”, providing concrete experimental evidence for integrating activation and blockade mechanisms within nanoplatforms for immunotherapy[194]. This highly controllable delivery system ensures spatiotemporal co-localization of multimodal chemoprophylactic therapy while minimizing circulating drug concentrations, avoiding immune-related adverse events (irAEs), and enhancing treatment safety and efficacy.
7.4 The role of exosomal PD-L1 and dynamic assessment of the TME
Exosomes are present in biological fluids such as blood and urine and can be used to non-invasively monitor the tumor immune microenvironment. Researchers have developed the NanoEPIC nanomagnetic sorting platform to address issues such as exosome heterogeneity and low detection sensitivity[219]. The platform uses magnetic nanoparticles to specifically label small extracellular vesicles and uses magnetic gradient sorting technology on a microfluidic chip to rapidly sort them based on PD-L1 expression levels. NanoEPIC scores showed a strong correlation between tumor volume and PD-L1 burden when observing the treatment window and efficacy. Clinical analysis results showed that patients who responded to immunotherapy had significantly lower exosomal PD-L1 levels, while patients who did not respond to immunotherapy had higher PD-L1 levels. This discovery provides an important biomarker for accurately identifying the treatment response window. In addition to static detection, real-time dynamic monitoring is also crucial for accurately identifying the treatment window. To this end, researchers have developed a microfluidic in vivo detection platform that integrates a perfusion system to continuously monitor fluctuations in the concentration of exosomes carrying PD-L1 in the blood[220]. Experimental results show that changes in PD-L1 concentration can accurately predict the time when tumors transition from slow growth to rapid expansion, thus laying a key foundation for real-time adjustment of clinical treatment plans. Furthermore, the discovery of PD-L1 in exosomes suggests its great potential for application in the precise molecular subtyping of tumors. Another researcher made a very sensitive chemiluminescence immunosensor out of nickel-cobalt hydroxide nanoflowers that can accurately measure the amount of PD-L1-positive exosomes in serum[221]. Clinical studies have shown that exosomal PD-L1 levels are closely associated with the invasiveness of lung adenocarcinoma. This platform can not only accurately diagnose lung adenocarcinoma, but it can also tell the difference between microinvasive and invasive adenocarcinoma. This platform can not only accurately identify lung adenocarcinoma with complete specificity, but also effectively distinguish between microinvasive adenocarcinoma and invasive adenocarcinoma. It can also serve as a reliable biomarker to assess the progression of the TME through liquid biopsy and to develop personalized treatment plans.
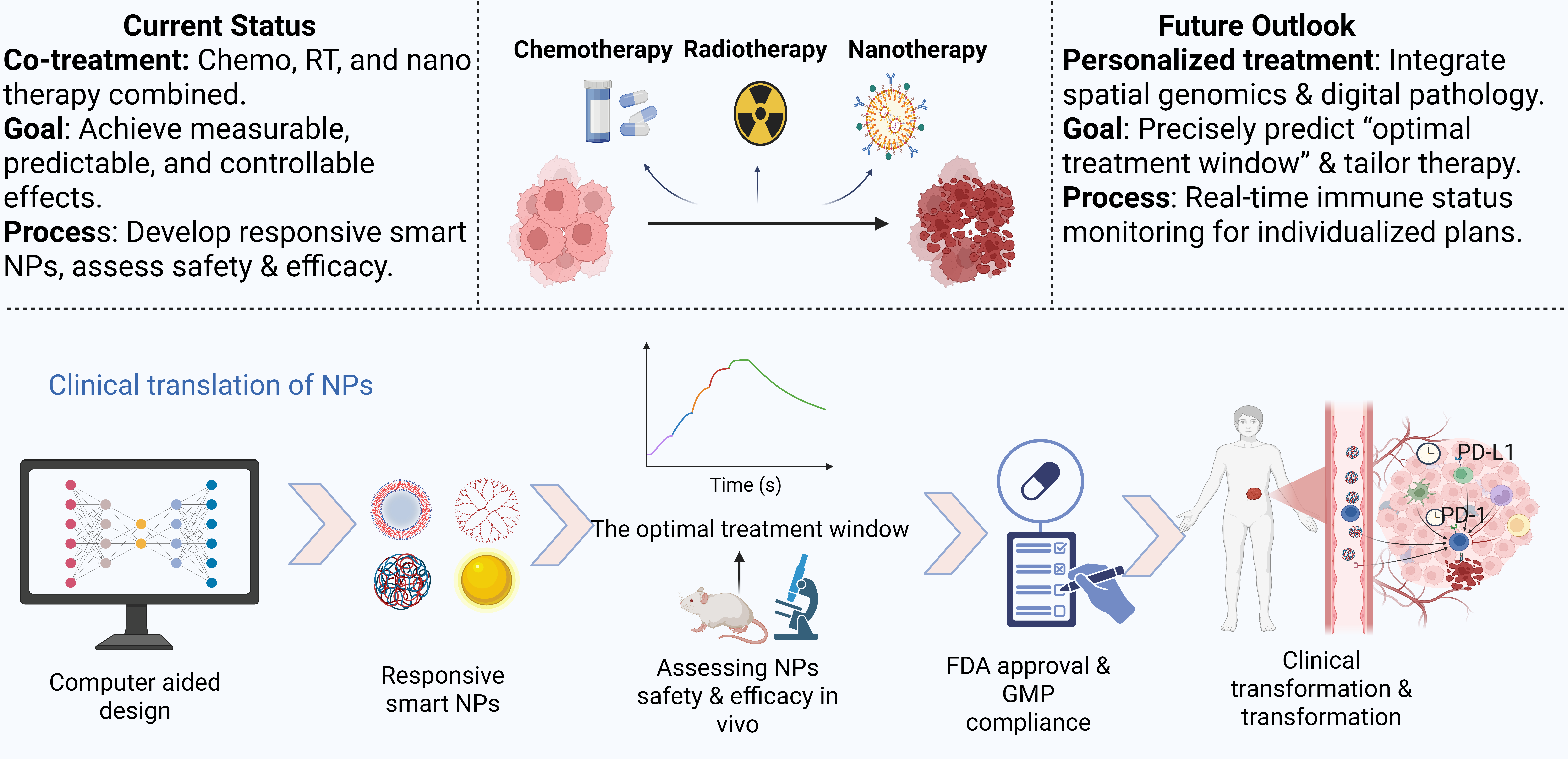
8. Clinical Translation and Future Outlook
In the next decade, sequential nanomedicine strategies for overcoming PD-1/PD-L1 resistance are expected to move from additive combinations toward more controllable and predictable treatment systems, where outcomes can be measured and adjusted in real time.
First, TME classification should move beyond qualitative descriptions and become a reproducible quantitative framework. This will require standardized immune features across patient groups, together with spatial omics and digital pathology to reduce bias from intratumoral heterogeneity.
Second, chemotherapy and RT induce time-dependent changes in PD-L1 expression, but these patterns differ across tumor types and treatment regimens. More longitudinal studies and modeling work are needed to define when PD-L1 and effector T cells peak, and how their timing differs. This will help make the “optimal intervention window” more precise and usable in practice.
In combined RT and immunotherapy, the cGAS-STING pathway and TREX1-related dose effects show that fractionation plays an important role in immune activation. Future strategies should therefore consider both radiation dose design and immune dynamics. At the same time, nanomedicine-based sequential delivery systems need to be tested in vivo to ensure that drug release matches the biological timing (Figure 11). For clinical use, these systems also need to meet basic manufacturing requirements such as consistent particle size, stable drug loading, and reproducible release profiles.
{kind=link}
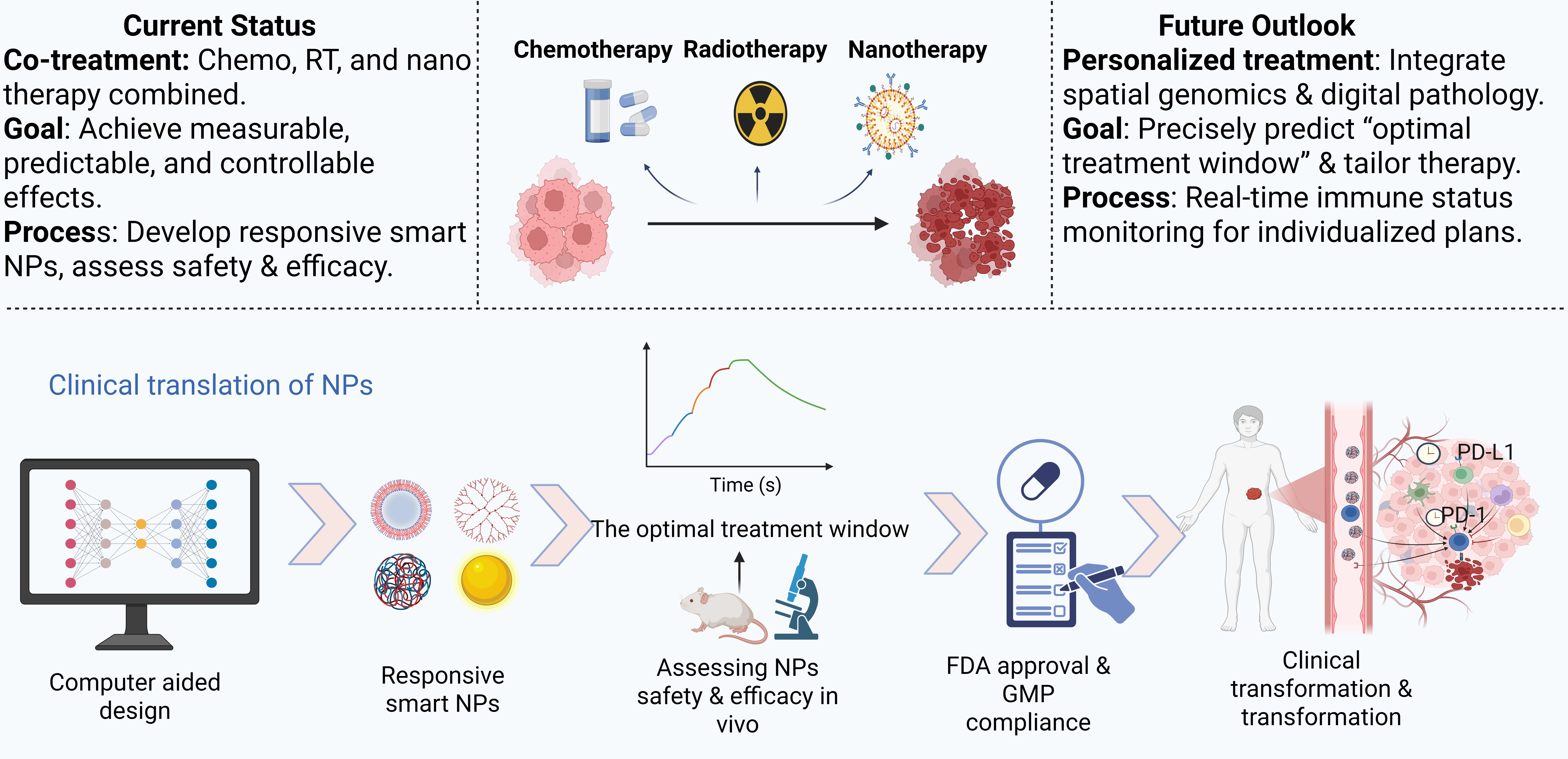
Figure 11. Schematic diagram illustrating the current status and future direction of nanomaterials in cancer treatment; currently, CRT and nanotherapy are combined, while in the future, personalized treatment and real-time immune monitoring will be used to optimize treatment outcomes. Created in BioRender. Lutf, B. (2026) https://BioRender.com/co4atey. CRT: chemoradiotherapy; RT: radiotherapy; NPs: nanoparticles.
Companion diagnostics will be essential for translating these strategies into practice. Tools such as exosomal PD-L1 detection, imaging, and microfluidic monitoring can help track immune status in real time. These markers should be integrated into clinical trials to guide patient selection, treatment timing, and early response evaluation. In the long term, a closed-loop system linking tumor subtype, timing window, drug delivery, monitoring, and treatment decisions may improve outcomes in patients who do not respond to current immunotherapy.
Despite encouraging progress in preclinical and translational studies, clinical evidence for sequential nanomedicine strategies remains limited. Most studies are still in early-stage clinical trials or preclinical models, and reliable randomized clinical data are scarce. Furthermore, as shown in Table 3, different nanoplatforms present specific safety concerns, including long-term storage stability of liposomes, off-target effects, and the risk of long-term bioaccumulation of nanomaterials, which are major obstacles to clinical application[47,222,223]. From a regulatory and CMC perspective, the complex structure of multifunctional nanoplatforms increases the difficulty of standardization, quality control, large-scale production, and approval processes, especially when multiple active ingredients and stimulus-response mechanisms are involved. Complex designs can reduce batch-to-batch consistency and reproducibility; therefore, future nanomedicine systems should be simpler, more modular, and easier to manufacture, while striking a balance between controlled delivery and therapeutic functionality.
Authors contribution
Lutf B: Conceptualization, writing-original draft.
Li Y: Writing-review & editing.
Shen J: Supervision, writing-review & editing.
Conflicts of interest
Jianliang Shen is an Editorial Board Member of BME Horizon. The other authors declare no conflicts of interest.
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Availability of data and materials
Not applicable.
Funding
None.
Copyright
© The Author(s) 2026.
References
-
1. Wu M, Huang Q, Xie Y, Wu X, Ma H, Zhang Y, et al. Improvement of the anticancer efficacy of PD-1/PD-L1 blockade via combination therapy and PD-L1 regulation. J Hematol Oncol. 2022;15(1):24.[DOI]
-
2. Sharma P, Goswami S, Raychaudhuri D, Siddiqui BA, Singh P, Nagarajan A, et al. Immune checkpoint therapy-current perspectives and future directions. Cell. 2023;186(8):1652-1669.[DOI]
-
3. Wang H, Chu Q, Ma S, Tao Y. Knockdown of programmed death 1 inhibited progression of papillary thyroid carcinoma in mice. Protein Pept Lett. 2023;30(5):396-400.[DOI]
-
4. Reck M, Rodríguez-Abreu D, Robinson AG, Hui R, Csőszi T, Fülöp A, et al. Pembrolizumab versus chemotherapy for PD-L1-positive non-small-cell lung cancer. N Engl J Med. 2016;375(19):1823-1833.[DOI]
-
5. Antonarakis ES, Piulats JM, Gross-Goupil M, Goh J, Ojamaa K, Hoimes CJ, et al. Pembrolizumab for treatment-refractory metastatic castration-resistant prostate cancer: Multicohort, open-label phase II KEYNOTE-199 study. J Clin Oncol. 2020;38(5):395-405.[DOI]
-
6. Le DT, Uram JN, Wang H, Bartlett BR, Kemberling H, Eyring AD, et al. PD-1 blockade in tumors with mismatch-repair deficiency. N Engl J Med. 2015;372(26):2509-20.[DOI]
-
7. Ju Y, Xu D, Liao MM, Sun Y, Bao WD, Yao F, et al. Barriers and opportunities in pancreatic cancer immunotherapy. NPJ Precis Oncol. 2024;8(1):199.[DOI]
-
8. Majidpoor J, Mortezaee K. The efficacy of PD-1/PD-L1 blockade in cold cancers and future perspectives. Clin Immunol. 2021;226:108707.[DOI]
-
9. McGrail DJ, Pilié PG, Rashid NU, Voorwerk L, Slagter M, Kok M, et al. High tumor mutation burden fails to predict immune checkpoint blockade response across all cancer types. Ann Oncol. 2021;32(5):661-672.[DOI]
-
10. Khosravi GR, Mostafavi S, Bastan S, Ebrahimi N, Gharibvand RS, Eskandari N. Immunologic tumor microenvironment modulators for turning cold tumors hot. Cancer Commun. 2024;44(5):521-553.[DOI]
-
11. Wu B, Zhang B, Li B, Wu H, Jiang M. Cold and hot tumors: From molecular mechanisms to targeted therapy. Sig Transduct Target Ther. 2024;9(1):274.[DOI]
-
12. Jiang Q, Chen Z, Jiang J, Chen Q, Lan H, Zhu J, et al. The role of cGAS-STING in remodeling the tumor immune microenvironment induced by radiotherapy. Crit Rev Oncol. 2025;209:104658.[DOI]
-
13. Antonangeli F, Natalini A, Garassino MC, Sica A, Santoni A, Di Rosa F et al. Regulation of PD-L1 expression by NF-κB in cancer. Front Immunol. 2020;11:584626.[DOI]
-
14. Chiang SF, Huang CY, Ke TW, Chen TW, Lan YC, You YS, et al. Upregulation of tumor PD-L1 by neoadjuvant chemoradiotherapy (neoCRT) confers improved survival in patients with lymph node metastasis of locally advanced rectal cancers. Cancer Immunol Immunother. 2019;68(2):283-296.[DOI]
-
15. Kciuk M, Kołat D, Kałuzińska-Kołat Ż, Gawrysiak M, Drozda R, Celik I, et al. PD-1/PD-L1 and DNA damage response in cancer. Cells. 2023;12(4):530.[DOI]
-
16. Yadollahi P, Jeon YK, Ng WL, Choi I. Current understanding of cancer-intrinsic PD-L1: Regulation of expression and its protumoral activity. BMB Rep. 2021;54(1):12-20.[DOI]
-
17. Cretella D, Digiacomo G, Giovannetti E, Cavazzoni A. PTEN alterations as a potential mechanism for tumor cell escape from PD-1/PD-L1 inhibition. Cancers. 2019;11(9):1318.[DOI]
-
18. Song M, Chen D, Lu B, Wang C, Zhang J, Huang L, et al. PTEN loss increases PD-L1 protein expression and affects the correlation between PD-L1 expression and clinical parameters in colorectal cancer. PLoS One. 2013;8(6):e65821.[DOI]
-
19. Zou J, Zhuang M, Yu X, Li N, Mao R, Wang Z, et al. MYC inhibition increases PD-L1 expression induced by IFN-γ in hepatocellular carcinoma cells. Mol Immunol. 2018;101:203-209.[DOI]
-
20. Wang NH, Lei Z, Yang HN, Tang Z, Yang MQ, Wang Y, et al. Radiation-induced PD-L1 expression in tumor and its microenvironment facilitates cancer-immune escape: A narrative review. Ann Transl Med. 2022;10(24):1406.[DOI]
-
21. Bae SU, Lee HW, Park JY, Seo I, Cho JM, Kim JY, et al. Neoadjuvant chemoradiotherapy up-regulates PD-L1 in radioresistant colorectal cancer. Clin Transl Radiat Oncol. 2025;51:100906.[DOI]
-
22. Zhang X, Cui H, Zhang X, Zhang Z, Liu Z, Mao R, et al. Chemotherapy modulates PD-L1 expression and its combination with immune checkpoint blockade requires precision. Crit Rev Oncol. 2025;214:104908.[DOI]
-
23. Wu CT, Chen WC, Chang YH, Lin WY, Chen MF. The role of PD-L1 in the radiation response and clinical outcome for bladder cancer. Sci Rep. 2016;6(1):19740.[DOI]
-
24. Yu Y, Zhang Q, Ma C, Yang X, Lin R, Zhang H, et al. Mesenchymal stem cells recruited by castration-induced inflammation activation accelerate prostate cancer hormone resistance via chemokine ligand 5 secretion. Stem Cell Res Ther. 2018;9(1):242.[DOI]
-
25. Wang W, Zhang S, Wang C, Gao S, Zhang L, Zhang C, et al. Fluorouracil enhances the anti-pancreatic cancer effect of anti-PD-L1 antibodies via up-regulating the expression of PD-L1 in cancer cells. J Investig Med. 2025;73(3):310-319.[DOI]
-
26. Makowska A, Meier S, Shen L, Busson P, Baloche V, Kontny U. Anti-PD-1 antibody increases NK cell cytotoxicity towards nasopharyngeal carcinoma cells in the context of chemotherapy-induced upregulation of PD-1 and PD-L1. Cancer Immunol Immunother. 2021;70(2):323-336.[DOI]
-
27. Sato H, Niimi A, Yasuhara T, Permata TBM, Hagiwara Y, Isono M, et al. DNA double-strand break repair pathway regulates PD-L1 expression in cancer cells. Nat Commun. 2017;8(1):1751.[DOI]
-
28. Volcic M, Karl S, Baumann B, Salles D, Daniel P, Fulda S, et al. NF-κB regulates DNA double-strand break repair in conjunction with BRCA1-CtIP complexes. Nucleic Acids Res. 2012;40(1):181-195.[DOI]
-
29. Storozynsky Q, Hitt MM. The impact of radiation-induced DNA damage on cGAS-STING-mediated immune responses to cancer. Int J Mol Sci. 2020;21(22):8877.[DOI]
-
30. Yue B, Gao W, Lovell JF, Jin H, Huang J. The cGAS-STING pathway in cancer immunity: Dual roles, therapeutic strategies, and clinical challenges. Essays Biochem. 2025;69(2):63-75.[DOI]
-
31. Singh AK, Winslow TB, Kermany MH, Goritz V, Heit L, Miller A, et al. A pilot study of stereotactic body radiation therapy combined with cytoreductive nephrectomy for metastatic renal cell carcinoma. Clin Cancer Res. 2017;23(17):5055-5065.[DOI]
-
32. Liu S, Wang W, Hu S, Jia B, Tuo B, Sun H, et al. Radiotherapy remodels the tumor microenvironment for enhancing immunotherapeutic sensitivity. Cell Death Dis. 2023;14(10):679.[DOI]
-
33. Gan Y, Li X, Han S, Liang Q, Ma X, Rong P, et al. The cGAS/STING pathway: A novel target for cancer therapy. Front Immunol. 2022;12:795401.[DOI]
-
34. Yang C, Liang Y, Liu N, Sun M. Role of the cGAS-STING pathway in radiotherapy for non-small cell lung cancer. Radiat Oncol. 2023;18(1):145.[DOI]
-
35. Padmanabhan S, Gaire B, Zou Y, Uddin MM, Vancurova I. IFNγ-induced PD-L1 expression in ovarian cancer cells is regulated by JAK1, STAT1 and IRF1 signaling. Cell Signal. 2022;97:110400.[DOI]
-
36. Sang H, Zhao X. JAK/STAT-mediated regulation of PD-L1 expression in breast cancer: Molecular mechanisms and implications for immunotherapy. iScience. 2026;29(3):114826.[DOI]
-
37. Basheeruddin M, Qausain S. Hypoxia-inducible factor 1-alpha (HIF-1α) and cancer: Mechanisms of tumor hypoxia and therapeutic targeting. Cureus. 2024;16(10):e70700.[DOI]
-
38. Triner D, Shah YM. Hypoxia-inducible factors: A central link between inflammation and cancer. J Clin Invest. 2016;126(10):3689-3698.[DOI]
-
39. Shurin MR, Umansky V. Cross-talk between HIF and PD-1/PD-L1 pathways in carcinogenesis and therapy. J Clin Invest. 2022;132(9):e159473.[DOI]
-
40. Yuan X, Liu X, Jiang D, Zheng Z, Ma X, Wu S, et al. Exosomal PD-L1 derived from hypoxia nasopharyngeal carcinoma cell exacerbates CD8+ T cell suppression by promoting PD-L1 upregulation in macrophages. Cancer Immunol Immunother. 2025;74(7):220.[DOI]
-
41. Bailey CM, Liu Y, Liu M, Du X, Devenport M, Zheng P, et al. Targeting HIF-1α abrogates PD-L1-mediated immune evasion in tumor microenvironment but promotes tolerance in normal tissues. J Clin Invest. 2022;132(9):e150846.[DOI]
-
42. Jin X, Zhang N, Yan T, Wei J, Hao L, Sun C, et al. Lactate-mediated metabolic reprogramming of tumor-associated macrophages: Implications for tumor progression and therapeutic potential. Front Immunol. 2025;16:1573039.[DOI]
-
43. Xanthopoulou ET, Kakouratos C, Nanos C, Gkegka AG, Kalaitzis C, Giatromanolaki A, et al. HIF1α-dependent and independent pathways regulate the expression of PD-L1 in prostate cancer. Med Oncol. 2023;40(5):151.[DOI]
-
44. Song CW, Kim H, Kim MS, Park HJ, Paek SH, Terezakis S, et al. Role of HIF-1α in the responses of tumors to radiotherapy and chemotherapy. Cancer Res Treat. 2025;57(1):1-10.[DOI]
-
45. Yu Y, Sun R, Hu F, Ding Z, Li X, Han J, et al. Hypoxia upregulates the expression of PD-L1 via NPM1 in breast cancer. J Immunother Cancer. 2025;13(6):e010151.[DOI]
-
46. Chen Z, Han F, Du Y, Shi H, Zhou W. Hypoxic microenvironment in cancer: Molecular mechanisms and therapeutic interventions. Sig Transduct Target Ther. 2023;8(1):70.[DOI]
-
47. Fan D, Cao Y, Cao M, Wang Y, Cao Y, Gong T. Nanomedicine in cancer therapy. Sig Transduct Target Ther. 2023;8(1):293.[DOI]
-
48. Peng J, Hamanishi J, Matsumura N, Abiko K, Murat K, Baba T, et al. Chemotherapy induces programmed cell death-ligand 1 overexpression via the nuclear factor-κB to foster an immunosuppressive tumor microenvironment in ovarian cancer. Cancer Res. 2015;75(23):5034-5045.[DOI]
-
49. Xiao Z, Su Z, Han S, Huang J, Lin L, Shuai X. Dual pH-sensitive nanodrug blocks PD-1 immune checkpoint and uses T cells to deliver NF-κB inhibitor for antitumor immunotherapy. Sci Adv. 2020;6(6):eaay7785.[DOI]
-
50. Sun M, Ni C, Li A, Liu J, Guo H, Xu F, et al. A biomimetic nanoplatform mediates hypoxia-adenosine axis disruption and PD-L1 knockout for enhanced MRI-guided chemodynamic-immunotherapy. Acta Biomater. 2025;201:618-632.[DOI]
-
51. Jiang D, Li Y. Unraveling the immunosuppressive microenvironment of glioblastoma and advancements in treatment. Front Immunol. 2025;16:1590781.[DOI]
-
52. Li D, Liu X, Gao W, Zhao W, Ji S, Wang S, et al. An immune subtype classification system enables the development of strategies to predict and enhance immunotherapy responses in colorectal cancer. Cancer Res. 2025;85(8):1441-1458.[DOI]
-
53. Wang L, Zhang L, Zhang Z, Wu P, Zhang Y, Chen X. Advances in targeting tumor microenvironment for immunotherapy. Front Immunol. 2024;15:1472772.[DOI]
-
54. Racacho KJ, Shiau YP, Villa R, Mahri S, Tang M, Lin TY, et al. The tumor immune microenvironment: Implications for cancer immunotherapy, treatment strategies, and monitoring approaches. Front Immunol. 2025;16:1621812.[DOI]
-
55. Vyhnánková S, Lacina L, Chovanec M, Plzák J, Smetana K Jr, Netušil J, et al. Cold, hot, and lethal-the tumour microenvironment and the immunology of head and neck squamous cell carcinoma. Int J Mol Sci. 2025;26(18):8844.[DOI]
-
56. Zheng S, Wang W, Shen L, Yao Y, Xia W, Ni C. Tumor battlefield within inflamed, excluded or desert immune phenotypes: The mechanisms and strategies. Exp Hematol Oncol. 2024;13(1):80.[DOI]
-
57. Liu YT, Wang YL, Wang S, Li JJ, He W, Fan XJ, et al. Turning cold tumors into hot tumors to ignite immunotherapy. Mol Cancer. 2025;24(1):254.[DOI]
-
58. Wang L, Geng H, Liu Y, Liu L, Chen Y, Wu F, et al. Hot and cold tumors: Immunological features and the therapeutic strategies. MedComm. 2023;4(5):e343.[DOI]
-
59. Dong Z, Yuan Z, Jin T, Gao C, Wang X, Xu F. Lactate at the crossroads of tumor metabolism and immune escape: A new frontier in cancer therapy. J Transl Med. 2025;23(1):1239.[DOI]
-
60. Zeng Y, Huang Y, Tan Q, Peng L, Wang J, Tong F, et al. Influence of lactate in resistance to anti-PD-1/PD-L1 therapy: Mechanisms and clinical applications (Review). Mol Med Rep. 2024;31(2):48.[DOI]
-
61. Zha J, Zhang J, Lu J, Zhang G, Hua M, Guo W, et al. A review of lactate-lactylation in malignancy: Its potential in immunotherapy. Front Immunol. 2024;15:1384948.[DOI]
-
62. Remley VA, Linden J, Bauer TW, Dimastromatteo J. Unlocking antitumor immunity with adenosine receptor blockers. Cancer Drug Resist. 2023;6(4):748-767.[DOI]
-
63. Bonaventura P, Shekarian T, Alcazer V, Valladeau-Guilemond J, Valsesia-Wittmann S, Amigorena S, et al. Cold tumors: A therapeutic challenge for immunotherapy. Front Immunol. 2019;10:168.[DOI]
-
64. Kwon WA, Joung JY. Immunotherapy in prostate cancer: From a “cold” tumor to a “hot” prospect. Cancers. 2025;17(7):1064.[DOI]
-
65. He C, Luo Y, Wang XL, Zhang C, Sun B. Leveraging immunogenic cell death in pancreatic cancer. Cancer Screen Prev. 2023;2(2):108-116.[DOI]
-
66. Wu Q, Xiao Q, Tang X, Li L, Song D, Zhou Y, et al. DAMPs prognostic signature predicts tumor immunotherapy, and identifies immunosuppressive mechanism of pannexin 1 channels in pancreatic ductal adenocarcinoma. Front Immunol. 2024;15:1516457.[DOI]
-
67. Jenkins L, Jungwirth U, Avgustinova A, Iravani M, Mills A, Haider S, et al. Cancer-associated fibroblasts suppress CD8+ T-cell infiltration and confer resistance to immune-checkpoint blockade. Cancer Res. 2022;82(16):2904-2917.[DOI]
-
68. Lv D, Fei Y, Chen H, Wang J, Han W, Cui B, et al. Crosstalk between T lymphocyte and extracellular matrix in tumor microenvironment. Front Immunol. 2024;15:1340702.[DOI]
-
69. Zhang C, Fei Y, Wang H, Hu S, Liu C, Hu R, et al. CAFs orchestrates tumor immune microenvironment-a new target in cancer therapy? Front Pharmacol. 2023;14:1113378.[DOI]
-
70. Zhang D, Song Q, Wang W, Li Q, Zhao Z, Jiang Y, et al. Unleashing a dual-warhead nanomedicine to precisely sensitize immunotherapy for pancreatic ductal adenocarcinoma. Adv Funct Mater. 2024;34(30):2315447.[DOI]
-
71. Bruni S, Mercogliano MF, Mauro FL, Cordo Russo RI, Schillaci R. Cancer immune exclusion: Breaking the barricade for a successful immunotherapy. Front Oncol. 2023;13:1135456.[DOI]
-
72. Hanley CJ, Thomas GJ. T-cell tumour exclusion and immunotherapy resistance: A role for CAF targeting. Br J Cancer. 2020;123(9):1353-1355.[DOI]
-
73. Kansha T, Ma X, Wang H, Yu X, Song Y, Guo Z, et al. Exosomal PD-L1 detection in cancer predictive biomarker for response to immune checkpoint blockade therapy. Front Immunol. 2025;16:1603855.[DOI]
-
74. Dragu LD, Chivu-Economescu M, Pitica IM, Matei L, Bleotu C, Diaconu CC, et al. Targeting exosomal PD-L1 as a new frontier in cancer immunotherapy. Curr Issues Mol Biol. 2025;47(7):525.[DOI]
-
75. Liu C, Liu D, Zheng X, Guan J, Zhou X, Zhang H, et al. Munc13-4 mediates tumor immune evasion by regulating the sorting and secretion of PD-L1 via exosomes. Nat Commun. 2025;16(1):9080.[DOI]
-
76. Tarin M, Oryani MA, Javid H, Karimi-Shahri M. Exosomal PD-L1 in non-small cell lung Cancer: Implications for immune evasion and resistance to immunotherapy. Int Immunopharmacol. 2025;155:114519.[DOI]
-
77. Wang B, Cheng D, Ma D, Chen R, Li D, Zhao W, et al. Mutual regulation of PD-L1 immunosuppression between tumor-associated macrophages and tumor cells: A critical role for exosomes. Cell Commun Signal. 2024;22(1):21.[DOI]
-
78. Shin K, Kim J, Park SJ, Lee MA, Park JM, Choi MG, et al. Prognostic value of soluble PD-L1 and exosomal PD-L1 in advanced gastric cancer patients receiving systemic chemotherapy. Sci Rep. 2023;13(1):6952.[DOI]
-
79. Chen J, Song Y, Miao F, Chen G, Zhu Y, Wu N, et al. PDL1-positive exosomes suppress antitumor immunity by inducing tumor-specific CD8+ T cell exhaustion during metastasis. Cancer Sci. 2021;112(9):3437-3454.[DOI]
-
80. Tufail M, Jiang CH, Li N. Immune evasion in cancer: Mechanisms and cutting-edge therapeutic approaches. Sig Transduct Target Ther. 2025;10(1):227.[DOI]
-
81. Lin P, Lin Y, Chen X, Zhao X, Cui L. Decoding MHC loss: Molecular mechanisms and implications for immune resistance in cancer. Clin Transl Med. 2025;15(7):e70403.[DOI]
-
82. Sari G, Rock KL. Tumor immune evasion through loss of MHC class-I antigen presentation. Curr Opin Immunol. 2023;83:102329.[DOI]
-
83. Wang J, Lu Q, Chen X, Aifantis I. Targeting MHC-I inhibitory pathways for cancer immunotherapy. Trends Immunol. 2024;45(3):177-187.[DOI]
-
84. Yu Q, Dong Y, Wang X, Su C, Zhang R, Xu W, et al. Pharmacological induction of MHC-I expression in tumor cells revitalizes T cell antitumor immunity. JCI Insight. 2024;9(17):e177788.[DOI]
-
85. Kong S, Zhang J, Wang L, Li W, Guo H, Weng Q, et al. Mechanisms of low MHC I expression and strategies for targeting MHC I with small molecules in cancer immunotherapy. Cancer Lett. 2025;611:217432.[DOI]
-
86. Taylor BC, Balko JM. Mechanisms of MHC-I downregulation and role in immunotherapy response. Front Immunol. 2022;13:844866.[DOI]
-
87. Sun X, Watanabe T, Oda Y, Shen W, Ahmad A, Ouda R, et al. Targeted demethylation and activation of NLRC5 augment cancer immunogenicity through MHC class I. Proc Natl Acad Sci U S A. 2024;121(6):e2310821121.[DOI]
-
88. Li C, Qi X, Yan M. Chemotherapy-induced immunogenic cell death in combination with ICIs: A brief review of mechanisms, clinical insights, and therapeutic implications. Front Pharmacol. 2025;16:1572195.[DOI]
-
89. Shi F, Huang X, Hong Z, Lu N, Huang X, Liu L, et al. Improvement strategy for immune checkpoint blockade: A focus on the combination with immunogenic cell death inducers. Cancer Lett. 2023;562:216167.[DOI]
-
90. Wang Y, Pattarayan D, Huang H, Zhao Y, Li S, Wang Y, et al. Systematic investigation of chemo-immunotherapy synergism to shift anti-PD-1 resistance in cancer. Nat Commun. 2024;15:[DOI]
-
91. Fumet JD, Limagne E, Thibaudin M, Ghiringhelli F. Immunogenic cell death and elimination of immunosuppressive cells: A double-edged sword of chemotherapy. Cancers. 2020;12(9):2637.[DOI]
-
92. Jeong H, Koh J, Kim S, Yim J, Song SG, Kim H, et al. Cell-intrinsic PD-L1 signaling drives immunosuppression by myeloid-derived suppressor cells through IL-6/Jak/Stat3 in PD-L1-high lung cancer. J Immunother Cancer. 2025;13(3):e010612.[DOI]
-
93. Chen P, Chen Z, Sui W, Han W. Recent advances in the mechanisms of PD-L1 expression in gastric cancer: A review. Biol Res. 2025;58(1):16.[DOI]
-
94. Ibrahim A, Mohamady Farouk Abdalsalam N, Liang Z, Kashaf Tariq H, Li R, O Afolabi L, et al. MDSC checkpoint blockade therapy: A new breakthrough point overcoming immunosuppression in cancer immunotherapy. Cancer Gene Ther. 2025;32(4):371-392.[DOI]
-
95. Jou E, Chaudhury N, Nasim F. Novel therapeutic strategies targeting myeloid-derived suppressor cell immunosuppressive mechanisms for cancer treatment. Explor Target Antitumor Ther. 2024;5(1):187-207.[DOI]
-
96. Lin X, Kang K, Chen P, Zeng Z, Li G, Xiong W, et al. Regulatory mechanisms of PD-1/PD-L1 in cancers. Mol Cancer. 2024;23(1):108.[DOI]
-
97. He Z, Jing X, Dai X, Bao L, Yang X, Xiong Y, et al. The ability of clinically relevant chemotherapeutics to induce immunogenic cell death in squamous cell carcinoma. Front Biosci. 2024;29(4):158.[DOI]
-
98. Li G, Che X, Wang S, Liu D, Xie D, Jiang B, et al. The role of cisplatin in modulating the tumor immune microenvironment and its combination therapy strategies: A new approach to enhance anti-tumor efficacy. Ann Med. 2025;57(1):2447403.[DOI]
-
99. Gandhi S, Slomba RT, Janes C, Fitzpatrick V, Miller J, Attwood K, et al. Systemic chemokine-modulatory regimen combined with neoadjuvant chemotherapy in patients with triple-negative breast cancer. J Immunother Cancer. 2024;12(11):e010058.[DOI]
-
100. Bracci L, Schiavoni G, Sistigu A, Belardelli F. Immune-based mechanisms of cytotoxic chemotherapy: Implications for the design of novel and rationale-based combined treatments against cancer. Cell Death Differ. 2014;21(1):15-25.[DOI]
-
101. Kim H, Chun SJ, Sun S, Cho H, Kim TJ, Lee YJ, et al. Low-dose cyclophosphamide enhances the tumoricidal effects of 5-day spacing stereotactic ablative radiotherapy by boosting antitumor immunity. Cancer Res Treat. 2025;57(3):678-692.[DOI]
-
102. Delahousse J, Molina L, Paci A. Cyclophosphamide and analogues; a matter of dose and schedule for dual anticancer activities. Cancer Lett. 2024;598:217119.[DOI]
-
103. Zitvogel L, Apetoh L, Ghiringhelli F, André F, Tesniere A, Kroemer G. The anticancer immune response: Indispensable for therapeutic success? J Clin Invest. 2008;118(6):1991-2001.[DOI]
-
104. Rinnerthaler G, Egle D, Bartsch R, Schmitt CA, Petzer A, Balic M, et al. Neoadjuvant atezolizumab in combination with dual HER2 blockade plus epirubicin in women with early HER2-positive breast cancer: The randomized phase 2 ABCSG-52/ATHENE trial. Nat Cancer. 2025;6(1):41-50.[DOI]
-
105. Wimmer K, Sachet M, Ramos C, Frantal S, Birnleitner H, Brostjan C, et al. Differential immunomodulatory effects of epirubicin/cyclophosphamide and docetaxel in breast cancer patients. J Exp Clin Cancer Res. 2023;42(1):300.[DOI]
-
106. Basak U, Sarkar T, Mukherjee S, Chakraborty S, Dutta A, Dutta S, et al. Tumor-associated macrophages: An effective player of the tumor microenvironment. Front Immunol. 2023;14:1295257.[DOI]
-
107. He Z, Zhang S. Tumor-associated macrophages and their functional transformation in the hypoxic tumor microenvironment. Front Immunol. 2021;12:741305.[DOI]
-
108. Zou Z, Lin H, Li M, Lin B. Tumor-associated macrophage polarization in the inflammatory tumor microenvironment. Front Oncol. 2023;13:1103149.[DOI]
-
109. Wu K, Lin K, Li X, Yuan X, Xu P, Ni P, et al. Redefining tumor-associated macrophage subpopulations and functions in the tumor microenvironment. Front Immunol. 2020;11:1731.[DOI]
-
110. Han S, Wang W, Wang S, Yang T, Zhang G, Wang D, et al. Tumor microenvironment remodeling and tumor therapy based on M2-like tumor associated macrophage-targeting nano-complexes. Theranostics. 2021;11(6):2892-2916.[DOI]
-
111. Liu S, Gong H, Li P, Hu J, Li Y, Xu R, et al. Chemotherapy-induced macrophage CXCL7 expression drives tumor chemoresistance via the STAT1/PHGDH-serine metabolism axis and SAM paracrine feedback to M2 polarization. Cell Death Dis. 2025;16(1):379.[DOI]
-
112. Hong Y, Fu Y, Long Q. Post-translational governance of NF-κB in cancer immunity: Mechanisms and therapeutic horizons. Front Immunol. 2025;16:1627084.[DOI]
-
113. Daniels MA, Teixeiro E. The NF-κB signaling network in the life of T cells. Front Immunol. 2025;16:1559494.[DOI]
-
114. Yi M, Zheng X, Niu M, Zhu S, Ge H, Wu K. Combination strategies with PD-1/PD-L1 blockade: Current advances and future directions. Mol Cancer. 2022;21(1):28.[DOI]
-
115. Feng X, Deng L, Gao C, Yang Z, Dong H, Xie S, et al. Hypoxia alleviation-potentiated chemotherapy enhances anti-PD-L1 treatment for triple-negative breast cancer via improving tumor immunogenicity and inhibiting STAT3 activation. ACS Appl Mater Interfaces. 2026;18(5):7845-7862.[DOI]
-
116. Jezierska-Drutel A, Rosenzweig SA, Neumann CA. Role of oxidative stress and the microenvironment in breast cancer development and progression. Adv Cancer Res. 2013;119:107-125.[DOI]
-
117. Li X, Wang S, Mu W, Barry J, Han A, Carpenter RL, et al. Reactive oxygen species reprogram macrophages to suppress antitumor immune response through the exosomal miR-155-5p/PD-L1 pathway. J Exp Clin Cancer Res. 2022;41(1):41.[DOI]
-
118. Krysko DV, Garg AD, Kaczmarek A, Krysko O, Agostinis P, Vandenabeele P. Immunogenic cell death and DAMPs in cancer therapy. Nat Rev Cancer. 2012;12(12):860-875.[DOI]
-
119. Teng Y, Gao L, Mäkitie AA, Florek E, Czarnywojtek A, Saba NF, et al. Iron, ferroptosis, and head and neck cancer. Int J Mol Sci. 2023;24(20):15127.[DOI]
-
120. Chung CH, Lin CY, Chen CY, Hsueh CW, Chang YW, Wang CC, et al. Ferroptosis signature shapes the immune profiles to enhance the response to immune checkpoint inhibitors in head and neck cancer. Adv Sci. 2023;10(15):e2204514.[DOI]
-
121. Roux C, Jafari SM, Shinde R, Duncan G, Cescon DW, Silvester J, et al. Reactive oxygen species modulate macrophage immunosuppressive phenotype through the up-regulation of PD-L1. Proc Natl Acad Sci U S A. 2019;116(10):4326-4335.[DOI]
-
122. Kang X, Zhang L, Liu S, Wang F, Liu H, Zhou F, et al. Platinum drugs upregulate CXCR4 and PD-L1 expression via ROS-dependent pathways, with implications for novel combined treatment in gastric cancer. J Pathol Clin Res. 2025;11(1):e70015.[DOI]
-
123. Wang D, Tang L, Zhang Y, Ge G, Jiang X, Mo Y, et al. Regulatory pathways and drugs associated with ferroptosis in tumors. Cell Death Dis. 2022;13(6):544.[DOI]
-
124. Calvani M, Subbiani A, Vignoli M, Favre C. Spotlight on ROS and β3-adrenoreceptors fighting in cancer cells. Oxid Med Cell Longev. 2019;2019:6346529.[DOI]
-
125. Jomova K, Alomar SY, Valko R, Fresser L, Nepovimova E, Kuca K, et al. Interplay of oxidative stress and antioxidant mechanisms in cancer development and progression. Arch Toxicol. 2026;100(1):27-73.[DOI]
-
126. Bianchi ME. Killing cancer cells, twice with one shot. Cell Death Differ. 2014;21(1):1-2.[DOI]
-
127. Arimoto KI, Miyauchi S, Liu M, Zhang DE. Emerging role of immunogenic cell death in cancer immunotherapy. Front Immunol. 2024;15:1390263.[DOI]
-
128. Catanzaro E, Beltrán-Visiedo M, Galluzzi L, Krysko DV. Immunogenicity of cell death and cancer immunotherapy with immune checkpoint inhibitors. Cell Mol Immunol. 2025;22(1):24-39.[DOI]
-
129. Janssens S, Rennen S, Agostinis P. Decoding immunogenic cell death from a dendritic cell perspective. Immunol Rev. 2024;321(1):350-370.[DOI]
-
130. Chen R, Zou J, Liu J, Kang R, Tang D. DAMPs in the immunogenicity of cell death. Mol Cell. 2025;85(20):3874-3889.[DOI]
-
131. Chen Y, Wang Z, Zhang C, Su Y, Zhou T, Hu K. Revealing the mechanism of natural product-induced immunogenic cell death: Opening a new chapter in tumor immunotherapy. Front Immunol. 2024;15:1470071.[DOI]
-
132. Han Y, Tian X, Zhai J, Zhang Z. Clinical application of immunogenic cell death inducers in cancer immunotherapy: Turning cold tumors hot. Front Cell Dev Biol. 2024;12:1363121.[DOI]
-
133. Calvillo-Rodríguez KM, Lorenzo-Anota HY, Rodríguez-Padilla C, Martínez-Torres AC, Scott-Algara D. Immunotherapies inducing immunogenic cell death in cancer: Insight of the innate immune system. Front Immunol. 2023;14:1294434.[DOI]
-
134. Esensten JH, Helou YA, Chopra G, Weiss A, Bluestone JA. CD28 costimulation: From mechanism to therapy. Immunity. 2016;44(5):973-988.[DOI]
-
135. Zanna MY, Yasmin AR, Omar AR, Arshad SS, Mariatulqabtiah AR, Nur-Fazila SH, et al. Review of dendritic cells, their role in clinical immunology, and distribution in various animal species. Int J Mol Sci. 2021;22(15):8044.[DOI]
-
136. Lu MM, Yang Y. Exosomal PD-L1 in cancer and other fields: Recent advances and perspectives. Front Immunol. 2024;15:1395332.[DOI]
-
137. Shareef A, Sameer HN, Yaseen A, Athab ZH, Adil M, Allela OQB. Recent advances in nanoparticles with a PD-L1-based therapy combination of chemotherapy for lung cancer. Crit Rev Oncol. 2025;214:104872.[DOI]
-
138. Fallatah MM, Alradwan I, Alfayez N, Aodah AH, Alkhrayef M, Majrashi M, et al. Nanoparticles for cancer immunotherapy: Innovations and challenges. Pharmaceuticals. 2025;18(8):1086.[DOI]
-
139. Ge Y, Zhou Q, Pan F, Wang R. Utilizing nanoparticles to overcome anti-PD-1/PD-L1 immunotherapy resistance in non-small cell lung cancer: A potential strategy. Int J Nanomedicine. 2025;20:2371-2394.[DOI]
-
140. Zhou S, Zhang Q, Han H, Chen G, Wang P, Xue S, et al. Hypoxia-activated paclitaxel prodrugs enable PD-L1 degradation to potentiate cancer chemo-immunotherapy. J Nanobiotechnology. 2025;24(1):18.[DOI]
-
141. Hu K, Zhang D, Ma W, Gu Y, Zhao J, Mu X. Polydopamine-based nanoparticles for synergistic chemotherapy of prostate cancer. Int J Nanomedicine. 2024;19:6717-6730.[DOI]
-
142. Liu D, Li K, Gong L, Fu L, Yang D. Charge reversal yolk-shell liposome co-loaded JQ1 and doxorubicin with high drug loading and optimal ratio for synergistically enhanced tumor chemo-immunotherapy via blockade PD-L1 pathway. Int J Pharm. 2023;635:122728.[DOI]
-
143. Deng L, Liang H, Xu M, Yang X, Burnette B, Arina A, et al. STING-dependent cytosolic DNA sensing promotes radiation-induced type I interferon-dependent antitumor immunity in immunogenic tumors. Immunity. 2014;41(5):843-852.[DOI]
-
144. Demaria S, Ng B, Devitt ML, Babb JS, Kawashima N, Liebes L, et al. Ionizing radiation inhibition of distant untreated tumors (abscopal effect) is immune mediated. Int J Radiat Oncol Biol Phys. 2004;58(3):862-870.[DOI]
-
145. Colangelo NW, Gerber NK, Vatner RE, Cooper BT. Harnessing the cGAS-STING pathway to potentiate radiation therapy: Current approaches and future directions. Front Pharmacol. 2024;15:1383000.[DOI]
-
146. Harding SM, Benci JL, Irianto J, Discher DE, Minn AJ, Greenberg RA. Mitotic progression following DNA damage enables pattern recognition within micronuclei. Nature. 2017;548(7668):466-470.[DOI]
-
147. Vanpouille-Box C, Alard A, Aryankalayil MJ, Sarfraz Y, Diamond JM, Schneider RJ, et al. DNA exonuclease Trex1 regulates radiotherapy-induced tumour immunogenicity. Nat Commun. 2017;8(1):15618.[DOI]
-
148. Ma C, Yu X, Zhang X, Su L, Jiang O, Cui R. Combination of radiotherapy and ICIs in advanced hepatocellular carcinoma: A systematic review of current evidence and future prospects (Review). Oncol Lett. 2025;30(1):342.[DOI]
-
149. Hovhannisyan L, Riether C, Aebersold DM, Medová M, Zimmer Y. CAR T cell-based immunotherapy and radiation therapy: Potential, promises and risks. Mol Cancer. 2023;22(1):82.[DOI]
-
150. Park G, Son B, Kang J, Lee S, Jeon J, Kim JH, et al. LDR-induced miR-30a and miR-30b target the PAI-1 pathway to control adverse effects of NSCLC radiotherapy. Mol Ther. 2019;27(2):342-354.[DOI]
-
151. He M, Chen S, Yu H, Fan X, Wu H, Wang Y, et al. Advances in nanoparticle-based radiotherapy for cancer treatment. iScience. 2025;28(1):111602.[DOI]
-
152. Wang B, Hu S, Teng Y, Chen J, Wang H, Xu Y, et al. Current advance of nanotechnology in diagnosis and treatment for malignant tumors. Sig Transduct Target Ther. 2024;9(1):200.[DOI]
-
153. Jackson N, Cecchi D, Beckham W, Chithrani DB. Application of high-Z nanoparticles to enhance current radiotherapy treatment. Molecules. 2024;29(11):2438.[DOI]
-
154. Zhang P, Darmon A, Marill J, Mohamed Anesary N, Paris S. Radiotherapy-activated hafnium oxide nanoparticles produce abscopal effect in a mouse colorectal cancer model. Int J Nanomedicine. 2020;15:3843-3850.[DOI]
-
155. Wang X, Wang D, Liao Y, Guo X, Song Q, Liu W, et al. Hafnium oxide-based sensitizer with radiation-triggered cuproptosis for radiotherapy. Nano Today. 2025;61:102626.[DOI]
-
156. Yi L, Jiang X, Zhou Z, Xiong W, Xue F, Liu Y, et al. A hybrid nanoadjuvant simultaneously depresses PD-L1/TGF-β1 and activates cGAS-STING pathway to overcome radio-immunotherapy resistance. Adv Mater. 2024;36(15):e2304328.[DOI]
-
157. Fucikova J, Kepp O, Kasikova L, Petroni G, Yamazaki T, Liu P, et al. Detection of immunogenic cell death and its relevance for cancer therapy. Cell Death Dis. 2020;11(11):1013.[DOI]
-
158. Shen M, Jiang X, Peng Q, Oyang L, Ren Z, Wang J, et al. The cGAS‒STING pathway in cancer immunity: Mechanisms, challenges, and therapeutic implications. J Hematol Oncol. 2025;18(1):40.[DOI]
-
159. Liu X, Yuan Z, Tang Z, Chen Q, Huang J, He L, et al. Selenium-driven enhancement of synergistic cancer chemo-/radiotherapy by targeting nanotherapeutics. Biomater Sci. 2021;9(13):4691-4700.[DOI]
-
160. Fan Y, Zhang J, Shi M, Li D, Lu C, Cao X, et al. Correction to “poly(amidoamine) dendrimer-coordinated copper(II) complexes as a theranostic nanoplatform for the radiotherapy-enhanced magnetic resonance imaging and chemotherapy of tumors and tumor metastasis”. Nano Lett. 2025;25(32):12436.[DOI]
-
161. Molinaro M, Skrodzki D, Pan D. Chemoradiotherapy and nanomedicine: Drug mechanisms and delivery systems. WIREs Nanomed Nanobiotechnol. 2024;16(4):e1984.[DOI]
-
162. Hagan CT, Mi Y, Knape NM, Wang AZ. Enhancing combined immunotherapy and radiotherapy through nanomedicine. Bioconjug Chem. 2020;31(12):2668-2678.[DOI]
-
163. Mousavi-Kiasary SMS, Senabreh A, Zandi A, Pena R, Cruz F, Adibi A, et al. Synergistic cancer therapies enhanced by nanoparticles: Advancing nanomedicine through multimodal strategies. Pharmaceutics. 2025;17(6):682.[DOI]
-
164. Johnstone TC, Suntharalingam K, Lippard SJ. The next generation of platinum drugs: Targeted Pt(II) agents, nanoparticle delivery, and Pt(IV) prodrugs. Chem Rev. 2016;116(5):3436-3486.[DOI]
-
165. Xiao X, Wang Y, Chen J, Qin P, Chen P, Zhou D, et al. Self-targeting platinum(IV) amphiphilic prodrug nano-assembly as radiosensitizer for synergistic and safe chemoradiotherapy of hepatocellular carcinoma. Biomaterials. 2022;289:121793.[DOI]
-
166. Soroushnasab H, Gharbavi M, Eskandari M, Rezaeejam H, Ghorbani R, Johari B. Methotrexate-conjugated silver nanoparticles enhance chemoradiotherapy effects on U87 human glioblastoma multiforme cells: As a radiosensitizer agent to improve X-irradiation efficacy. J Drug Deliv Sci Technol. 2025;107:106791.[DOI]
-
167. Su L, Zhu K, Ge X, Wu Y, Zhang J, Wang G, et al. X-ray activated nanoprodrug for visualization of cortical microvascular alterations and NIR-II image-guided chemo-radiotherapy of glioblastoma. Nano Lett. 2024;24(12):3727-3736.[DOI]
-
168. Yao W, Zhao X, Gong Y, Zhang M, Zhang L, Wu Q, et al. Impact of the combined timing of PD-1/PD-L1 inhibitors and chemotherapy on the outcomes in patients with refractory lung cancer. ESMO Open. 2021;6(2):100094.[DOI]
-
169. Huo Y, Wang D, Yang S, Xu Y, Qin G, Zhao C, et al. Optimal timing of anti-PD-1 antibody combined with chemotherapy administration in patients with NSCLC. J Immunother Cancer. 2024;12(12):e009627.[DOI]
-
170. Lee SS, Pagacz J, Averbek S, Scholten D, Liu Y, Kron SJ. Timing anti-PD-L1 checkpoint blockade immunotherapy to enhance tumor irradiation. Cancers. 2025;17(3):391.[DOI]
-
171. Yi H, Luo Q, Bai Z, Li X, Yu Y, Shi J, et al. Precise combination therapy of radiotherapy and immunotherapy guided by joint PD-L1 imaging and T-cell imaging. J Immunother Cancer. 2025;13(11):e012300.[DOI]
-
172. Pouw JEE, Hashemi SMS, Huisman MC, Wijngaarden JE, Slebe M, Oprea-Lager DE, et al. First exploration of the on-treatment changes in tumor and organ uptake of a radiolabeled anti PD-L1 antibody during chemoradiotherapy in patients with non-small cell lung cancer using whole body PET. J Immunother Cancer. 2024;12(2):e007659.[DOI]
-
173. Zhong L, Peng W, Sun J, Luo Y, Sheng H, Wu Y, et al. Impact of timing the combination of radiotherapy and PD-1 inhibitors on outcomes in patients with hepatocellular carcinoma. J Hepatocell Carcinoma. 2025;12:123-134.[DOI]
-
174. Landré T, Karaboué A, Buchwald ZS, Innominato PF, Qian DC, Assié JB, et al. Effect of immunotherapy-infusion time of day on survival of patients with advanced cancers: A study-level meta-analysis. ESMO Open. 2024;9(2):102220.[DOI]
-
175. Xu Z, Liu S, Li Y, Wu Y, Tu J, Chen Q, et al. Engineering strategies of sequential drug delivery systems for combination tumor immunotherapy. Acta Pharm Sin B. 2025;15(8):3951-3977.[DOI]
-
176. Wang Y, Wang Z, Chen B, Yin Q, Pan M, Xia H, et al. Cooperative self-assembled nanoparticle induces sequential immunogenic cell death and toll-like receptor activation for synergistic chemo-immunotherapy. Nano Lett. 2021;21(10):4371-4380.[DOI]
-
177. Reda M, Ngamcherdtrakul W, Nelson MA, Siriwon N, Wang R, Zaidan HY, et al. Development of a nanoparticle-based immunotherapy targeting PD-L1 and PLK1 for lung cancer treatment. Nat Commun. 2022;13(1):4261.[DOI]
-
178. Zhang T, Tang D, Wu P, Jiang S, Zhang Y, Naeem A, et al. NIR-II photo-accelerated polymer nanoparticles boost tumor immunotherapy via PD-L1 silencing and immunogenic cell death. Bioact Mater. 2025;46:285-300.[DOI]
-
179. Lian S, Yang W, Zeng Y, Tang R, Wang K. Targeted nano-drug delivery systems for tumor immunotherapy. J Pharm Anal. 2026;16(1):101408.[DOI]
-
180. Zhang X, Wang X, Hou L, Xu Z, Liu Y, Wang X. Nanoparticles overcome adaptive immune resistance and enhance immunotherapy via targeting tumor microenvironment in lung cancer. Front Pharmacol. 2023;14:1130937.[DOI]
-
181. Huynh TMH, Huang PX, Wang KL, Tran NT, Iao HM, Pan WC, et al. Reprogramming immunodeficiency in lung metastases via PD-L1 siRNA delivery and antigen capture of nanosponge-mediated dendritic cell modulation. ACS Nano. 2025;19(27):25134-25153.[DOI]
-
182. Kowal M, Woźniacka R, Ścisłowska-Czarnecka A, Homa J, Głodzik W. The effect of physical activity on cytokine levels in adults living with type 1 diabetes-a preliminary study. Physiol Rep. 2024;12(8):e15985.[DOI]
-
183. Zhou Z, Li C, Li C, Zhou L, Tan S, Hou W, et al. Mitochondria-targeted nanoadjuvants induced multi-functional immune-microenvironment remodeling to sensitize tumor radio-immunotherapy. Adv Sci. 2024;11(26):e2400297.[DOI]
-
184. Hu R, Jiang X, Zhu L, Meng R, Yang R, Sun W, et al. Overcoming radiation-induced PD-L1 and COX-2 upregulation by nitric oxide gas nanogenerator to sensitize radiotherapy of lung cancer. Biomaterials. 2025;321:123335.[DOI]
-
185. Xu J, Li Z, Tong Q, Zhang S, Fang J, Wu A, et al. CD133+PD-L1+ cancer cells confer resistance to adoptively transferred engineered macrophage-based therapy in melanoma. Nat Commun. 2025;16(1):895.[DOI]
-
186. Teng Y, Luo C, Qiu X, Mu J, Sriwastva MK, Xu Q, et al. Plant-nanoparticles enhance anti-PD-L1 efficacy by shaping human commensal microbiota metabolites. Nat Commun. 2025;16(1):1295.[DOI]
-
187. Hernandez-Franco JF, Jan IM, Elzey BD, HogenEsch H. Intradermal vaccination with a phytoglycogen nanoparticle and STING agonist induces cytotoxic T lymphocyte-mediated antitumor immunity. NPJ Vaccines. 2024;9(1):149.[DOI]
-
188. Cai X, Bao X, Wu Y. Metal-organic frameworks as intelligent drug nanocarriers for cancer therapy. Pharmaceutics. 2022;14(12):2641.[DOI]
-
189. Gharehbaba AM, Omidi Y, Barar J, Eskandani M, Adibkia K. Synergistic pH-responsive MUC-1 aptamer-conjugated Ag/MSN Janus nanoparticles for targeted chemotherapy, photothermal therapy, and gene therapy in breast cancer. Biomater Adv. 2025;166:214081.[DOI]
-
190. Wang Z, Tang XL, Zhao MJ, Zhang YD, Xiao Y, Liu YY, et al. Biomimetic hypoxia-triggered RNAi nanomedicine for synergistically mediating chemo/radiotherapy of glioblastoma. J Nanobiotechnology. 2023;21(1):210.[DOI]
-
191. Hu S, Huang L, Zhou L, Wu T, Zhao S, Zhang L. Single-excitation triple-emission down-/up-conversion nanoassemblies for tumor microenvironment-enhanced ratiometric NIR-II fluorescence imaging and chemo-/photodynamic combination therapy. Anal Chem. 2023;95(7):3830-3839.[DOI]
-
192. Deng S, Hu L, Chen G, Ye J, Xiao Z, Guan T, et al. A PD-L1 siRNA-loaded boron nanoparticle for targeted cancer radiotherapy and immunotherapy. Adv Mater. 2025;37(13):e2419418.[DOI]
-
193. Barati M, Mirzavi F, Nikpoor AR, Sankian M, Namdar Ahmadabad H, Soleimani A, et al. Enhanced antitumor immune response in melanoma tumor model by anti-PD-1 small interference RNA encapsulated in nanoliposomes. Cancer Gene Ther. 2022;29(6):814-824.[DOI]
-
194. Fu Y, Huang Y, Li P, Wang L, Tang Z, Liu X, et al. Physical- and chemical-dually ROS-responsive nano-in-gel platforms with sequential release of OX40 agonist and PD-1 inhibitor for augmented combination immunotherapy. Nano Lett. 2023;23(4):1424-1434.[DOI]
-
195. Lee Y, Song S, Yang S, Kim J, Moon Y, Shim N, et al. Photo-induced crosslinked and anti-PD-L1 peptide incorporated liposomes to promote PD-L1 multivalent binding for effective immune checkpoint blockade therapy. Acta Pharm Sin B. 2024;14(3):1428-1440.[DOI]
-
196. Park HB, An EK, Kim SJ, Ryu D, Zhang W, Pack CG, et al. Anti-PD-L1 antibody fragment linked to tumor-targeting lipid nanoparticle can eliminate cancer and its metastasis via photoimmunotherapy. ACS Nano. 2024;18(49):33366-33380.[DOI]
-
197. Xiao C, Wang X, Li S, Zhang Z, Li J, Deng Q, et al. A cuproptosis-based nanomedicine suppresses triple negative breast cancers by regulating tumor microenvironment and eliminating cancer stem cells. Biomaterials. 2025;313:122763.[DOI]
-
198. Ouyang P, Wang L, Wu J, Tian Y, Chen C, Li D, et al. Overcoming cold tumors: A combination strategy of immune checkpoint inhibitors. Front Immunol. 2024;15:1344272.[DOI]
-
199. Liu YT, Sun ZJ. Turning cold tumors into hot tumors by improving T-cell infiltration. Theranostics. 2021;11(11):5365-5386.[DOI]
-
200. Ying W, Zheng C, Gong C, Yuan J, Xu G, Zhou J, et al. Nanorobots hold PD-L1 and break membrane of colorectal cancer cells for immunotherapy. Nat Nanotechnol. 2026;21(1):156-167.[DOI]
-
201. Zhou Z, Luo W, Zheng C, Wang H, Hu R, Deng H, et al. Mitochondrial metabolism blockade nanoadjuvant reversed immune-resistance microenvironment to sensitize albumin-bound paclitaxel-based chemo-immunotherapy. Acta Pharm Sin B. 2024;14(9):4087-4101.[DOI]
-
203. Soko GF, Kosgei BK, Meena SS, Ng YJ, Liang H, Zhang B, et al. Extracellular matrix re-normalization to improve cold tumor penetration by oncolytic viruses. Front Immunol. 2024;15:1535647.[DOI]
-
204. Provenzano PP, Cuevas C, Chang AE, Goel VK, von Hoff DD, Hingorani SR, et al. Enzymatic targeting of the stroma ablates physical barriers to treatment of pancreatic ductal adenocarcinoma. Cancer Cell. 2012;21(3):418-429.[DOI]
-
205. Gordon-Weeks A, Yuzhalin AE. Cancer extracellular matrix proteins regulate tumour immunity. Cancers. 2020;12(11):3331.[DOI]
-
206. Shabbir F, Mujeeb AA, Jawed SF, Khan AH, Shakeel CS. Simulation of transvascular transport of nanoparticles in tumor microenvironments for drug delivery applications. Sci Rep. 2024;14(1):1764.[DOI]
-
207. Yonucu S, Yιlmaz D, Phipps C, Unlu MB, Kohandel M. Quantifying the effects of antiangiogenic and chemotherapy drug combinations on drug delivery and treatment efficacy. PLoS Comput Biol. 2017;13(9):e1005724.[DOI]
-
208. Zhou Z, Jiang X, Yi L, Li C, Wang H, Xiong W, et al. Mitochondria energy metabolism depression as novel adjuvant to sensitize radiotherapy and inhibit radiation induced-pulmonary fibrosis. Adv Sci. 2024;11(26):e2401394.[DOI]
-
209. Castiglioni A, Yang Y, Williams K, Gogineni A, Lane RS, Wang AW, et al. Combined PD-L1/TGFβ blockade allows expansion and differentiation of stem cell-like CD8 T cells in immune excluded tumors. Nat Commun. 2023;14(1):4703.[DOI]
-
210. Liu M, Ma X, Zhou R, Yang X, Zhou Y, Ma B, et al. Bioengineered anti-PD-L1 functionalized nanoplatform for targeted delivery and tumor immune reprogramming against colorectal cancer. Biomater Res. 2025;29:0284.[DOI]
-
211. Li C, Xiong W, Liu J, Li K, Wang H, Wang Z, et al. Nanoparticles-mediated mitochondrial relocation of lipid-lowering drugs shape energy metabolism to conquer acquired immune resistance. Drug Resist Updat. 2026;84:101323.[DOI]
-
212. Katebi A, Riazi-Rad F, Ahangari F, Ajdary S. M1 macrophage membrane-engineered PLGA nanoparticles reprogram M2 tumor-associated macrophages to enhance anti-tumor immunity in breast cancer. J Mater Sci Mater Med. 2026;37(1):41.[DOI]
-
213. Xia S, Chen W, Xu Z, Gao Y, Chen J, Ding N, et al. Targeting Dicer reprograms tumor-associated macrophages to promote anti-tumoral immunity in colorectal cancer liver metastasis. J Nanobiotechnology. 2025;23(1):421.[DOI]
-
214. Zhang X, Liu Y, Jiang M, Mas-Rosario JA, Fedeli S, Cao-Milan R, et al. Polarization of macrophages to an anti-cancer phenotype through in situ uncaging of a TLR 7/8 agonist using bioorthogonal nanozymes. Chem Sci. 2024;15(7):2486-2494.[DOI]
-
215. Kim H, Khanna V, Kucaba TA, Zhang W, Ferguson DM, Griffith TS, et al. Combination of sunitinib and PD-L1 blockade enhances anticancer efficacy of TLR7/8 agonist-based nanovaccine. Mol Pharm. 2019;16(3):1200-1210.[DOI]
-
216. Su Q, Wang Z, Li P, Wei X, Xiao J, Duan X. pH and ROS dual-responsive autocatalytic release system potentiates immunotherapy of colorectal cancer. Adv Healthc Mater. 2024;13(29):e2401126.[DOI]
-
217. Huang Y, Wu S, Li J, He C, Cheng Y, Li N, et al. Self-amplified pH/ROS dual-responsive co-delivery nano-system with chemo-photodynamic combination therapy in hepatic carcinoma treatment. Int J Nanomedicine. 2024;19:3737-3751.[DOI]
-
218. Zou J, Jiang C, Hu Q, Jia X, Wang S, Wan S, et al. Tumor microenvironment-responsive engineered hybrid nanomedicine for photodynamic-immunotherapy via multi-pronged amplification of reactive oxygen species. Nat Commun. 2025;16(1):424.[DOI]
-
219. Chen K, Duong BTV, Ahmed SU, Dhavarasa P, Wang Z, Labib M, et al. A magneto-activated nanoscale cytometry platform for molecular profiling of small extracellular vesicles. Nat Commun. 2023;14(1):5576.[DOI]
-
220. Cong XZ, Feng J, Zhang HJ, Zhang LZ, Lin TY, Chen G, et al. Microfluidic device-based in vivo detection of PD-L1-positive small extracellular vesicles and its application for tumor monitoring. Anal Chem. 2024;96(6):2658-2665.[DOI]
-
221. Wang M, Shu J, Wang Y, Zhang W, Zheng K, Zhou S, et al. Ultrasensitive PD-L1-expressing exosome immunosensors based on a chemiluminescent nickel-cobalt hydroxide nanoflower for diagnosis and classification of lung adenocarcinoma. ACS Sens. 2024;9(6):3444-3454.[DOI]
-
222. Havelikar U, Ghorpade KB, Kumar A, Patel A, Singh M, Banjare N, et al. Comprehensive insights into mechanism of nanotoxicity, assessment methods and regulatory challenges of nanomedicines. Discov Nano. 2024;19(1):165.[DOI]
-
223. Souto EB, Blanco-Llamero C, Krambeck K, Kiran NS, Yashaswini C, Postwala H, et al. Regulatory insights into nanomedicine and gene vaccine innovation: Safety assessment, challenges, and regulatory perspectives. Acta Biomater. 2024;180:1-17.[DOI]
Copyright
© The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
Publisher’s Note
Science Exploration remains a neutral stance on jurisdictional claims in published
maps
and institutional affiliations. The views expressed in this article are solely those
of
the author(s) and do not reflect the opinions of the Editors or the publisher.
Share And Cite



Science Exploration Style
Lutf B, Li Y, Shen J. Nanomaterial-based optimization of chemo-radiotherapy for precisely overcoming PD-L1 resistance in the tumor immune microenvironment. BME Horiz. 2026;4:202617. https://doi.org/10.70401/bmeh.2026.0031
Tips
Copy completed.
Submit a Manuscript
Author Instructions
Cite this Article
Article Metrics
0
View
0
Download
Cited
Article Updates

- Abstract
- Keywords
- 1. Introduction
- 2. Molecular Mechanisms of PD-L1 Resistance and Heterogeneity of TME
- 3. The Immunomodulatory Role of Chemotherapy in TME Remodeling
- 4. Local Effects of RT
- 5. The Mechanism of Synergy Between Chemotherapy and Radiation Therapy
- 6. Finding the Optimal Intervention Window for Time-Series Strategies
- 7. Multimodal Embedding and Spatiotemporal Controlled Release in Nanotechnology
- 8. Clinical Translation and Future Outlook
- Authors contribution
- Conflicts of interest
- Ethical approval
- Consent to participate
- Consent for publication
- Availability of data and materials
- Funding
- References
- Copyright
Science Exploration Style
Lutf B, Li Y, Shen J. Nanomaterial-based optimization of chemo-radiotherapy for precisely overcoming PD-L1 resistance in the tumor immune microenvironment. BME Horiz. 2026;4:202617. https://doi.org/10.70401/bmeh.2026.0031
copy
Share Link
copy